Woda Budowa i właściwości
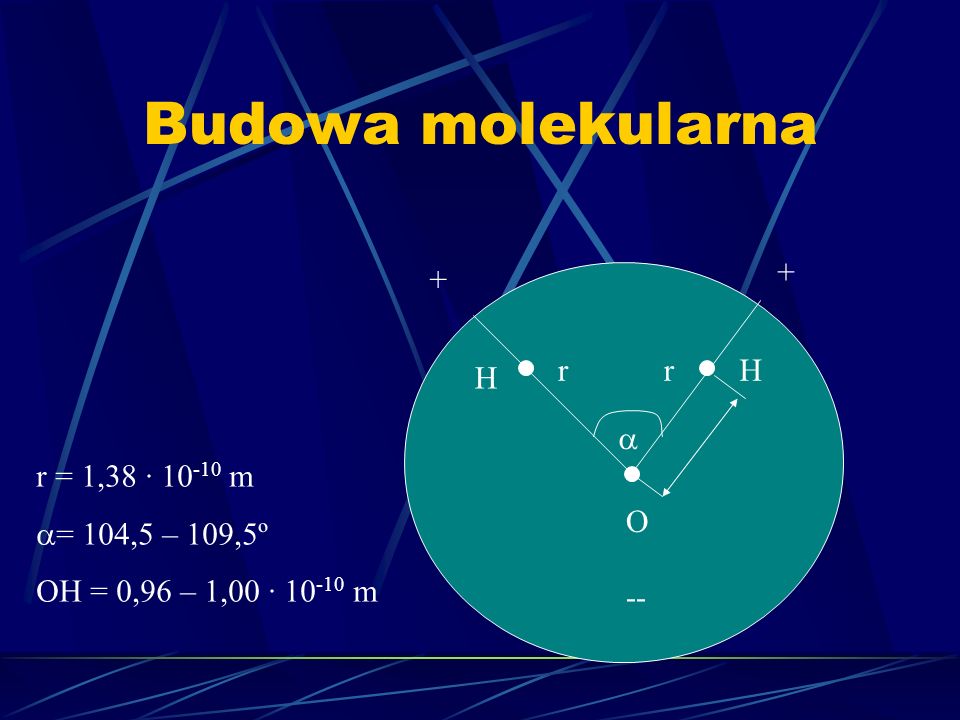
Budowa molekularna + + r r H H r = 1,38 · 10-10 m = 104,5 – 109,5º OH = 0,96 – 1,00 · 10-10 m O --
Charakterystyka cząsteczki wody Kowalencyjne wiązania między atomami tworzącymi cząsteczkę Kowalencyjne wiązania między cząsteczkami wody gdzie tlen jest akceptorem elektronu a grupa OH – donorem; ich liczba zależy od stanu skupienia wody (lód zawiera 4) Posiada właściwości polarne wynikające z rozmieszczenia ładunków, a jej moment dipolowy wynosi = 0.61 · 10-27 C · m
Struktura tetraedryczna lodu Charakterystyka: Każdy wierzchołek tetraedru zawiera cząsteczkę wody Odległość między atomami tlenu wynosi 2,76 · 10-10 m Odległości miedzy tlenem i wodorem tej samej cząsteczki wynoszą 0,99 · 10-10 m Odległości miedzy tlenem i wodorem sąsiednich cząsteczek wynoszą 1,77 · 10-10 m
Struktura krystaliczna lodu Zależy od ciśnienia i temperatury W normalnych warunkach gęstość lodu wynosi 920 kg · m-3 a strukturę tworzą pierścienie sześcioczłonowe ułożone w pomarszczone warstwy
Struktura krystaliczna lodu Każda cząsteczka otoczona przez 4 inne (3 w tej samej warstwie i 1 w następnej), Układ cząsteczek tworzy regularny czworościan w którym poszczególne tetraedry oddalone są o 4,5 · 10-10 m Atomy wodoru oscylują wokół atomów tlenu w odległości 0,96 · 10-10 m Nie jest strukturą sztywną ze względu na duży udział wolnej przestrzeni (ok. 66%) Duży udział wolnej przestrzeni decyduje o kruchości i porowatości lodu.
Charakterystyka wody w stanie ciekłym Nie jest dokładnie poznana (dotyczy to zwłaszcza wody obecnej w układach biologicznych) Zachowuje część wiązań wodorowych We współczesnych modelach zakłada się występowanie obszarów nieuporządkowanych i uporządkowanych kulistych (klatrat) lub pentagonalnych
Charakterystyka wody w stanie ciekłym Energia potrzebna do zerwania wiązań wodorowych w lodzie (ciepło topnienia) = 333 kJ · kg-1 Ta sama energia w przypadku wody ciekłej (ciepło parowania) = 2260 kJ · kg-1 Wartości te wskazują, że podczas topnienia lodu zrywanych jest tylko 15% wiązań wodorowych
Charakterystyka wody w stanie ciekłym Elementy klaretowe: składają się z 20 cząsteczek rozłożonych równomiernie po powierzchni kuli, przypomina piłkę o bokach z 12 regularnych pięcioboków. W jej wnętrzu znajduje się związana cza steczka wody lub inna. Odległości między atomami większe niż w strukturze lodu, Elementy pentagonalne: zawierają na krawędziach czworoboki ze zwiniętymi narożnikami. Struktura pośrednia miedzy klaretem i lodem.
Zmiany właściwości wody w obecności makrocząsteczek Zmiany temperatury powodują nieliniowe zmiany parametrów wody, tj. gęstość, lepkość, napięcie powierzchniowe, ciepło właściwe i ciepło parowania na skutek zmian struktury cząsteczki Bipolimery, za pośrednictwem powierzchni granicznych, stabilizują struktury cząsteczki wody w czasie i przestrzeni Połączenia między wodą a biopolimerami zmieniają się z częstotliwością 1011 s-1. Ich przykładem są wiązania wodorowe pomiędzy wodą a grupami aminowymi i karboksylowymi białek, które powodują zmiany struktury w białkach
Biofizyka makrocząsteczek c.d. Kwasy nukleinowe Biofizyka makrocząsteczek c.d.
Kwasy nukleinowe (NA)
Lokalizacja DNA w jądrze komórkowym
Połączenia DNA i histonów w jądrze komórkowym
Funkcje biologiczne Makrocząsteczki o właściwościach polinukleotydów i wysokiej masie cząsteczkowej,
Funkcje biologiczne Makrocząsteczki o właściwościach polinukleotydów i wysokiej masie cząsteczkowej, Tworzą strukturę genów i bezpośrednio biorą udział w biosyntezie białka.
Właściwości chemiczne Ze względu na udział 3 atomów H w cząsteczce H3PO4 i reszty pentozy tworzy jednozasadową strukturę diestrową, pK reakcji jonizacji wynosi ok. 1,0, co decyduje o stabilności przy niskich wartościach pH, Mają skłonność do łatwego tworzenia soli – nukleinianów, powstających z udziałem anionu nukleinowego, Reszty cukrowcowe łączą się z heterocyklicznymi zasadami azotowymi za pośrednictwem wiązań N-glikozydowych
Właściwości chemiczne Pentozy występują w formie -furanozowej, Ryboza w gorących kwasach przekształca się w furfural, co stanowi podstawę reakcji orcynolowej, Deoksyryboza w podobnym środowisku tworzy kwas -hydroksylewulinowy, który reaguje z difenyloaminą, Ze względu na wiązania N-glikozydowe kwasy nukleinowe nie ulegają hydrolizie zasadowej, a jedynie kwaśnej (w przypadku RNA hydrolizę zasadową umożliwia obecność grupy 2’-hydroksylowej)
Jednostki strukturalne Polinukleotydy (kwasy nukleinowe)
Jednostki strukturalne Oligonukleotydy (fragmenty złożone z kilku nukleotydów)
Jednostki strukturalne Nukleotydy (rybotydy, deoksy- rybotydy) Nukleozydy (rybozydy, deoksyry- bozydy)
Zasady azotowe (aminy drugorzędowe) Purynowe Pirymidynowe
Pentozy
Pentozy
Kwas fosforowy (fosforan) H3PO4
Komplementarność zasad według Watson’a i Crick’a
Składniki nietypowe 2’metylo-ryboza, Zasady metylowane, acetylowane, zredukowane i modyfikowane na inne sposoby, Pseudourydyna (nukleozyd, w którym U łączy się z rybozą wiązaniem C-G’ Oligosacharydowe łańcuchy boczne w DNA wirusowych, Rzadkie zasady i nukleozydy: ksantyna (Xan) i ksantozyna (Xao), hipoksantyna (Hyp), inozyna (Ino), kwas orotowy i orotydyna (Oro i Ord)
Składniki nietypowe Cukry inne niż ryboza, których obecność zaznacza się stosując przed skrótem nazwy nukleozydu lub nukleotydu odpowiednią małą literę, np. dThd, 3’-dGMP, 5’-dGMP d – deoksyryboza, a – arabinoza, x – ksyloza, l - liksoza
Składniki nietypowe (nukleozydy)
Zasady zapisu sekwencji kwasu nukleinowego Natura cukru dla RNA i DNA wynika jedynie z zapisu, Dla identyfikacji poszczególnych nukleotydów stosuje się pojedyncze litery (A, G, C, U, T), Obecność grupy fosforanowej zaznacza się znakiem łącznika, Modyfikacje zasad oznacza się za pomocą małych liter umieszczonych przed dużymi lub indeksów Modyfikacji cukrów zapisuje się za pomocą małych liter po symbolach nukleozydów
Zapis sekwencji kwasów nukleinowych - U – 5’UMP U - – 3’UMP A - U – fosforan dwunukleozydowy bez wolnej grupy fosforanowej A - U - – fosforan dwunukleozydowy z wolną grupą fosforanową przy końcu 3’ - A - U – fosforan dwunukleozydowy z wolną grupą fosforanową przy końcu 5’
Zapis sekwencji kwasów nukleinowych c.d. A – C – G – T – G – A – T - fragment łańcucha kwasu nukleinowego zawierający grupę –OH przy końcu 5’ i grupę fosforanową przy końcu 3’ A p C p G p T p G p A p T p Modyfikacja powyższego zapisu polegająca na zamianie znaku „-” literą „p” oznaczającą grupę fosforanową ppA – ADP; pppA - ATP
Zapis sekwencji kwasów nukleinowych c.d. A – U! - wykrzyknik oznacza obecność cyklicznej grupy 2’, 3’ fosforanowej przy urydynie Elementy niezidentyfikowane: Zasada purynowa: R lub Ph Zasada pirymidynowa Y lub Py Nukleotyd N
Zapis sekwencji kwasów nukleinowych c.d. Skróty oznaczające pochodne: ac – octanowe m – metylowe h – uwodornione s – tiopochodne (pochodne siarczkowe)
Zapis sekwencji kwasów nukleinowych c.d. C – hU – A – m62A – I – ac4C – Gm – G – OH5’ – cytydyno – 5,6 – dihydroksyurydyno – adenozyno – 6 – dimetyloadenozyno – inozyno – 4 – acetylocytydyno – 2 – metyloguanozyno – guanozyna.
Struktura wtórna kwasów nukleinowych konformacja cząsteczki Forma przestrzenna, którą przyjmują cząsteczki w wyniku wszystkich oddziaływań wewnątrz cząsteczkowych, które mogą być dodatkowo modyfikowane przez czynniki zewnętrzne
Struktura wtórna DNA (James Watson & Maurice Crick1953)
Struktura wtórna DNA (1953)
Struktura wtórna kwasów nukleinowych Struktura bihelix zaproponowana przez J.Watsona i M.Cricka w 1953 r. Odkrycie to poprzedziły: badania rentgenograficzne Rosalind Franklin, dotyczące kwasów strąconych z roztworów wodnych poprzez dodanie rozpuszczalników organicznych, Opracowanie reguł Pero Chargraffa, na podstawie stosunku zasad azotowych w kwasach nukleinowych
Reguły Chargraffa (1950) 1. [A] = [T] 1. [G] = [C]
Reguły Chagraffa 1. A + C = G + T 2. G + A = C + T 3. 4. A + T + G + C = constans
Stosunek zasad azotowych w kwasach nukleinowych
Struktura drugorzędowa kwasów nukleinowych W oparciu o reguły Chargraffa i metody pośrednie, stwierdzono że G + C = 38%, co daje G = 19% i C = 19%; A + T = 62%, co daje z kolei A = 31% i T = 31%
Struktura drugorzędowa kwasów nukleinowych Analiza rentgenostrukturalna wykonana przez Rosalind Franklin wskazywała na istnienie w cząsteczce DNA konformacji helikalnej
Rentgenogram DNA
Nadbudowanie zasad Zasady sąsiadujące ze sobą w heliksie, pomimo iż nie są komplementarne, orientują się w sposób równoległy względem cząsteczek sąsiednich, Oddziaływanie to jest stosunkowo silne i umożliwia wykrywanie wpływu określonych zasad daleko od ich lokalizacji.
Struktura drugorzędowa DNA – klasyczny model Watsona i Cricka
Struktura drugorzędowa DNA – klasyczny model Watsona i Cricka
Kierunki analizy konformacji kwasów nukleinowych Analiza konformacji nukleotydów, prowadzona metodami: - spektroskopowyni, - rentgenostrukturalnymi, - innymi metodami optycznymi
Kierunki analizy konformacji kwasów nukleinowych Analiza teoretyczna – modelowanie cząsteczek, uwzględniające możliwe układy stereodynamiczne i energetyczne, zgodnie z zasadą, że stabilnym (optymalnym) konformacjom muszą towarzyszyć minimalne wartości energii
Analiza rentgenostrukturalna kwasów nukleinowych Pełna analiza konformacyjna jest niemożliwa, ze względu na to, że kwasy nukleinowe nie występują w postaci krystalicznej,
Analiza rentgenostrukturalna kwasów nukleinowych Pełna analiza konformacyjna jest niemożliwa, ze względu na to, że kwasy nukleinowe nie występują w postaci krystalicznej, Jedynie reszty pentozowe i fosforanowe tworzą prawidłową siatkę przestrzenną, co stwarza możliwość analizy osi heliksu pod kątem liczby ekwiwalentnych grup występujących w jednym skręcie
Metody badań kwasów nukleinowych Spektrofotometria w nadfiolecie (UV) i podczerwieni (IR), Pomiary właściwości termodynamicznych, Dyspersja skręcania płaszczyzny polaryzacji, Widma dichroizmu kołowego (CD), Pomiary dwójłomności w przepływie, Badania hydrodynamiczne, Techniki spektrometryczne NMR.
Metody badań kwasów nukleinowych c.d. Pomiary dichroizmu kołowego umożliwiły ocenę wpływu wilgotności względnej i stężenia jonów na zmiany konformacji kwasów nukleinowych,
Metody badań kwasów nukleinowych c.d. Spektrometria w podczerwieni (IR) nukleozydów, nukleotydów i kwasów nukleinowych, wykorzystuje drgania, charakterystyczne dla poszczególnych ugrupowań wchodzących w skład kwasów nukleinowych
Pasma (cm-1) charakterystycznych drgań w kwasach nukleinowych 2800-3500 OH NH CH woda, cukier reszty zasad cukier i reszty zasad 1800-1500 C = O C = N C = C vNH vHOH } reszty zasad woda Ok. 1200 PO- antysymetryczne walencyjne - drgania walencyjne, v – drgania płaskie deformujące, - drgania deformujące niepłaskie
Pasma (cm-1) charakterystycznych drgań w kwasach nukleinowych 1100-1000 PO2 CO symetryczne walencyjne cukier 1000-700 PO NH grupa fosforanowa reszty zasad 600-300 deformacyjne drgania wszystkich grup - drgania walencyjne, v – drgania płaskie deformujące, - drgania deformujące niepłaskie
Metody badań kwasów nukleinowych c.d. Spektrometria w podczerwieni (IR) dowiodła istotnej roli wody w stabilizacji konformacji DNA. Przy wilgotności 92% 1 nukleotyd DNA jest uwodniony przez 20 cząsteczek wody, co jest niezbędne do utrzymania regularnej struktury heliksu.
Metody badań kwasów nukleinowych c.d. Widma nukleotydów w nadfiolecie (UV)
Układ przestrzenny osi cząsteczki kwasu nukleinowego Kierunek łańcucha 11 kątów rotacji wokół wiązań Zasada
Parametry najczęściej brane pod uwagę przy analizach konformacji kwasów nukleinowych 1. Główne: - długość wiązań walencyjnych, - kąty wartościowości, - kąty dwuścienne jednostek monomerycznych tworzących heliks
Parametry najczęściej brane pod uwagę przy analizach konformacji kwasów nukleinowych c.d. 2. Pomocnicze: - 1 - kąt obrotu płaszczyzny zasady azotowej wokół osi y’ (1), wiążącej dwa atomy wiąznia glikozydowego C1, - 2 - kąt obrotu wokół osi x’ – prostopadłej do osi heliksu (przechodzącej przez C4 pirymidyn i C8 puryn), - D – odległość początku liniowego układu współrzędnych od osi heliksu,
Parametry najczęściej brane pod uwagę przy analizach konformacji kwasów nukleinowych c.d. 2. Pomocnicze: - d – translacja wzdłuż osi heliksu d = c/n, gdzie c – okres identyczności n – liczba par zasad w okresie, - - kąt obrotu wokół osi heliksu = 2/n
Rodzaje DNA A-DNA Z-DNA B-DNA
Formy B- i A-DNA
Charakterystyka B-DNA Heliks prawoskrętny Pary zasad ułożone równolegle względem siebie i prostopadle do osi heliksu w odstępach 0,34nm, Skok o długości 3,4nm zawiera 10 par zasad, Atomy P występują w odległości 0,9nm od osi heliksu, Liczba zasad w obu komplementarnych heliksach może być różna
Charakterystyka A-DNA Heliks prawoskrętny, Pary zasad nachylone pod kątem 20° względem ich ułożenia w B-DNA, Na jeden skok heliksu przypada 11 par zasad, W skrajnych wartościach pH i temperatury może ulegać denaturacji, Heliksy mogą różnić się liczbą zasad.
Zmiany właściwości optycznych towarzyszące Denaturacji DNA Zmiana konformacji cząsteczki, Wzrost absorbancji (zwłaszcza przy 260nm), Wzrost energii swobodnej Gibbsa (G).
Zmiany właściwości optycznych towarzyszące Denaturacji DNA
Forma Z-DNA Oś heliksu Zasady
Charaktrystyka Z-DNA Heliks lewoskrętny, Forma zbliżona do B-DNA, Posiada 9,3 par zasad na 1 skok heliksu
Charakterystyka T-DNA Występuje u bakteriofagów T2, T4 i T6 przy wilgotności 66-44%, Zawiera 8 par zasad w jednym skoku heliksu, Kąt obrotu sąsiednich par zasad wokół osi heliksu jest największy ze wszystkich znanych rodzajów DNA i wynosi 42 - 45°
Inne rodzaje dwuniciowego DNA C-DNA – heliks prawoskrętny, powstaje we włóknach przy wilgotności względnej 57-66% i posiada 9,3 par zasad na obrót helisy, D-DNA – heliks prawoskrętny, występuje w odcinakach poli(dA)·poli(dT) i zawiera 8,5 par zasad na obrót helisy
Trój- i czteroniciowe formy DNA
H-DNA
Wiązania wodorowe w H-DNA Para zasad Watsona-Cricka Para zasad Hoogsteena
Wiązania wodorowe w trójniciowych NA c.d.
Rodziny kwasów nukleinowych Rodzina A A-DNA wszystkie helikalne rodzaje RNA Rodzina B B-DNA Z-DNA T-DNA
Wpływ jonów i wilgotności względnej na konformację DNA Formą najczęściej występującą w warunkach naturalnych jest B-DNA, A-DNA występuje w obecności jonów Na+ i przy wilgotności względnej wynoszącej 75%, B-DNA występuje przy 92% wilgotności względnej, a w obecności jonów Li+ nawet przy 66%, Z-DNA, w obecności jonów Li+ przy 44% wilgotności względnej.
Szereg przejściowy BT Formy przejściowe DNA Szereg przejściowy BT obejmujący formy zawierające 8; 8,33; 9; 9,33 par zasad na jeden skok. wzrost kąta między parami zasad od 36° do 45° spadek skoku od 3,37 do 2,72 nm i średnicy cząsteczki z 2,55 do 2,15 nm. Przejście AB wymaga znacznych zmian w środowisku zewnętrznym, ma charakter kooperatywny i jest związane ze zmianą konformacją furanozy.
Kwas rybonukleinowy RNA
Konformacje RNA
Formy bihelikalne RNA na ogół zawiera 11 par zasad na skok heliksu (forma A-RNA), w obecności NaCl przechodzi w formę A’-RNA (AA’), zawierającą 12 par zasad na skok, A’’-RNA, najmniej poznana, zwiera 11,5 nukleotydów na jeden skok heliksu
Rodzaje RNA Cytoplazmatyczny, Jądrowy
Cytoplazmatyczny RNA rRNA tRNA mRNA
r – RNA 1. Stanowi większość komórkowego RNA, 2 r – RNA 1. Stanowi większość komórkowego RNA, 2. Jest składnikiem rybosomów, stanowiąc 50-60% masy 3. Masa cząsteczkowa uzależnina od jednostki rybosomu i rodzaju komórki; podjednostki mniejsze 5501-7502 tys. Da, większe 1.11 do 1.52 mln Da 1Prokaryota 2Eukaryota
t-RNA
t-RNA Zawiera 75 – 85 nukleotydów w jednym łańcuchu, który w pewnych fragmentach staje się komplementarny, „Na dole cząsteczki” zawiera antykodon, oddziaływujący z odpowiadającym kodonem w kierunku antyrónoległym. Trzeci nukleozyd kodonu ma prawdopodobnie zdolność tworzenia wiązań wodorowych zgodnie z regułą Wooble (G-U, I-C, I-U, I-A), co umożliwia 1 cząsteczce t-RNA rozpoznawanie kilku kodonów kodujących dany aminokwas,
Różne typy wiązań wodorowych w cząsteczkach kwasów nukleinowych
t-RNA Cechy wspólne t-RNA: występowanie pętli T-pseudourydylo-C (T-Ψ-C) „po prawej stronie” ugrupowanie DU (dihydrourydyna) „po stronie lewej”, sekwencja końca 3’ G-C-A oraz w większości cząsteczek występowanie na końcu 5’ nukleozydu G.
Przykłady t-RNA
Przykłady t-RNA
Charakterystyka m-RNA Masa cząsteczkowa od 25000 Da do kilku milionów, Ogromne zróżnicowanie sekwencji nukleotydowej, Oprócz zasadniczej części kodującej sekwencję aminokwasów w konkretnym białku może zawierać sekwencje niekodujące, Może występować w kompleksach z białkami zwłaszcza w komórkach eukariotycznych
Budowa m-RNA Począwszy od końca 5’ mRNA składa się z następujących odcinków: Czapeczka, zbudowana z 7-metylo-Gppp i kolejnych dwóch zmetylowanych nukleotydów, Lider, zbudowany z 50-80 nukleotydów, Odcinek kodujący jeden łańcuch polipeptydowy, Odcinek poliA zbudowany z 50-200 reszt kwasu adenylowego Uwaga: w mRNA prokariota nie występuje poliA; kwas ten zawiera prawie wyłącznie sekwencje kodujące kilka białek
Funkcje biologiczne mRNA – kod genetyczny składający się z 61 kodonów sensownych i 3 niesensowne
Charakterystyka cytoplazmatycznych rodzajów RNA
Cytoplazmatyczny RNA hnRNA snRNA
Heterogenny jądrowy RNA (hnRNA) Prekursor cytoplazmatycznych form RNA, Cząsteczki niestabilne, Zawierają sekwencje nieobecne w cytoplazmatycznych frakcjach RNA – introny, Przekształcenie hnRNA następuje w procesie składania RNA, obejmującym wycinanie intronów, W procesie tym bierze udział wiele małych jądrowych RNA – snRNA, szczegółnie szeroko rozpowszechnionych w eukariotycznych komórkach ssaków
U1 snRNA człowieka
Struktura czwartorzędowa kwasów nukleinmowych Połączenia z innymi kwasami nukleinowymi Połączenia z białkami
Połączenia z kwasami nukleinowymi
Połączenia z białkami (w jądrze)
Połączenia z białkami (w cząsteczkach wirusów)
(w cząsteczkach wirusów) Połączenia z białkami (w cząsteczkach wirusów)