Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
Cykl komórkowy

2
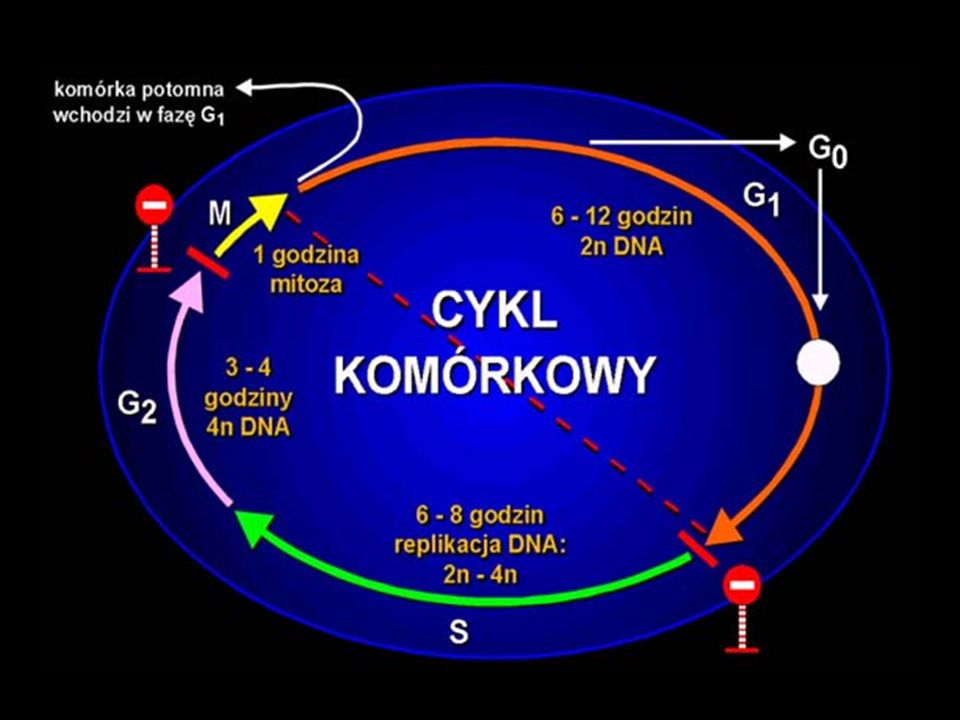
Cykl komórkowy jest szeregiem zmian biofizycznych i biochemicznych komórki, zachodzących między końcem jednego i końcem następnego podziału. Jeśli cykl komórkowy kończy się mitozą, wówczas określa się go mianem cyklu mitotycznego. Cykl komórkowy składa się z interfazy, czyli okresu pomiędzy dwoma podziałami komórki oraz samego podziału czyli mitozy. W interfazie zachodzi zjawisko replikacji materiału genetycznego.

3
W czasie mitozy podwojony materiał genetyczny jest rozdzielony w równych częściach do dwóch potomnych komórek. Replikacje materiału genetycznego i podział komórki zachodzące w cyklu komórkowym są podstawą zachowania ciągłości cech między kolejnymi pokoleniami komórek. W interfazie cyklu komórkowego wyróżnia się fazę G1 (ang. gap – przerwa) między końcem mitozy, a rozpoczęciem syntezy DNA, fazę syntezy (faza S) oraz fazę G2 między końcem syntezy DNA, a początkiem mitozy.
 między końcem mitozy, a rozpoczęciem syntezy DNA, fazę syntezy (faza S) oraz fazę G2 między końcem syntezy DNA, a początkiem mitozy.")
5
Interfaza

6
centrosom błona jądrowa jąderko chromatyna cytozol błona komórkowa Interfaza

7
Faza G1 Jest okresem życia komórki od końca mitozy, do rozpoczęcia syntezy DNA. Komórka wchodząca w tę fazę jest dwukrotnie mniejsza niż jej komórka matka. W fazie G1 wyróżnia się kilka stadiów oznaczonych jako: G1A, G1B, G1C itp. W zależności od stopnia syntezy oraz zawartości RNA i białek trwają one różnie długo, nawet w tej samej populacji komórek.

8
Faza G1 Faza G1 charakteryzuje się intensywnymi procesami anabolicznymi, znacznym stopniem wymian chemicznych z otoczeniem, oraz wzrostem innych przejawów aktywności jak: ruchliwość, pinocytoza, transport przez błony itp. We wczesnej fazie G1 komórka osiąga punkt restrykcyjny (punkt R) – jeśli go przekroczy, wówczas podejmuje syntezę DNA i kończy cykl podziałem. Jeśli zaś nie może przekroczyć punktu R, wówczas wchodzi w fazę spoczynkową (faza G0).
 – jeśli go przekroczy, wówczas podejmuje syntezę DNA i kończy cykl podziałem. Jeśli zaś nie może przekroczyć punktu R, wówczas wchodzi w fazę spoczynkową (faza G0).")
9
Faza S Przed każdym podziałem mitotycznym komórki ilość DNA przypadająca na jądro podwaja się, aby każda z dwóch potomnych komórek mogła otrzymać właściwą część DNA. Podwojenie ilości DNA dokonuje się w ograniczonym czasie interfazy zwanym fazą syntezy (S) DNA. Synteza DNA jest zaprogramowana genetycznie i nazwano ją programowaną syntezą DNA, która przebiega według sposobu semikonserwatywnego, tzn. podwójna spirala DNA ulega rozdzieleniu, a na każdej z 2 nici syntetyzowana jest nowa nić.
 DNA. Synteza DNA jest zaprogramowana genetycznie i nazwano ją programowaną syntezą DNA, która przebiega według sposobu semikonserwatywnego, tzn. podwójna spirala DNA ulega rozdzieleniu, a na każdej z 2 nici syntetyzowana jest nowa nić.")
10
Faza G2 cyklu komórkowego - między zakończeniem fazy S, a rozpoczęciem mitozy. Trwa zazwyczaj kilka godzin. Istotnym procesem zachodzącym w tej fazie jest synteza części białek wrzeciona podziałowego, głównie tubuliny. W fazie G2 odbywa się także intensywna produkcja składników potrzebnych do odtwarzania błon otoczki jądrowej i plazmolemmy komórek w telofazie i cytokinezie. Pod koniec fazy G2 następuje uaktywnienie kompleksu p34 – cyklina (kinaza fazy H), co inicjuje kaskadę fosforylacji i defosforylacji białek.
, co inicjuje kaskadę fosforylacji i defosforylacji białek.")
11
Faza G0 Faza G0 jest stanem spoczynkowym komórki – komórki funkcjonują, jednak tracą zdolność odtwarzania materiału genetycznego i dzielenia się. Czas trwania tej fazy jest różnie długi – od kilku dni, do kilku miesięcy i dłużej. Pod wpływem różnych bodźców komórki mogą z fazy G0 wchodzić z powrotem do fazy G1 cyklu komórkowego. Im dłużej komórki pozostają w fazie G0 tym więcej czasu zabiera im wejście w cykl po pobudzeniu.

12
Wchodzenie w cykl komórkowy
Do wejścia komórki w cykl prowadzi uruchomienie programu redukcji (programu plejotypowego). Składają się na niego w kolejności pojawiania się: - synteza hnRNA i rRNA – 3-12 godzin od chwili zadziałania bodźca - synteza białek enzymatycznych i strukturalnych – godzin od zadziałania bodźca - synteza DNA i histonów – godzin od zadziałania bodźca - synteza białek wrzeciona podziałowego i mitoza – godziny od zadziałania bodźca.
. Składają się na niego w kolejności pojawiania się: - synteza hnRNA i rRNA – 3-12 godzin od chwili zadziałania bodźca. - synteza białek enzymatycznych i strukturalnych – godzin od zadziałania bodźca. - synteza DNA i histonów – godzin od zadziałania bodźca. - synteza białek wrzeciona podziałowego i mitoza – godziny od zadziałania bodźca.")
13
Mitoza Jest formą zwiększania liczby komórek przez podział. Można w niej wyróżnić: - kariokinezę czyli podział jądra - profaza - prometafaza - metafaza - anafaza - telofaza - cytokinezę czyli podział cytoplazmy

14
Kariokineza

15
Profaza i prometafaza stają się widoczne chromosomy, które powstały z silnie skręconej chromatyny chromosomy składają się z dwóch połówek - chromatyd zanika jąderko na terenie cytoplazmy organizuje się wrzeciono podziałowe (w komórkach zwierzęcych uczestniczą w tym centriole) zanika błona jądrowa
 zanika błona jądrowa.")
16
błona jądrowa chromosomy jąderko cytozol błona komórkowa centromer chromosom = 2 siostrzane chromatydy połączone centromerem Wczesna profaza

17
fragmenty błony jądrowej
mikrotubule centrosom cytozol błona komórkowa centromer chromosom = 2 siostrzane chromatydy połączone centromerem Późna profaza

18
Metafaza uwolnione z jądra komórkowego chromosomy przemieszczają się w rejon równika wrzeciona podziałowego centromery leżą w płaszczyźnie równika, a w tym miejscu do chromosomów przyczepione są włókna wrzeciona centromery pękają

19
płytka metafazalna (równikowa)
centrosom cytozol błona komórkowa centromer chromosom = 2 siostrzane chromatydy połączone centromerem Metafaza

20
Anafaza włókna wrzeciona kurczą się
chromatydy - od tego momentu chromosomy potomne, wędrują ku biegunom wrzeciona podziałowego

21
centrosom cytozol błona komórkowa chromosom = 2 siostrzane chromatydy połączone centromerem Wczesna anafaza

22
bruzda podziałaowa chromosom błona komórkowa Późna anafaza

23
Telofaza chromosomy potomne rozkręcają się tworząc znów chromatynę
odtwarzają się jąderka wokół każdego ze skupień chromatyny odtwarza się błona jądrowa

24
bruzda podziałaowa Telofaza

25
Cytokineza

26
Podział cytoplazmy, który prowadzi do powstania dwóch komórek potomnych. Zaczyna się pod koniec anafazy lub na początku telofazy powstaniem pierścienia kurczliwego, który jest nagromadzeniem filamentów aktynowych i miozynowych pod błoną komórkową w płaszczyźnie prostopadłej do długiej osi wrzeciona podziałowego. Pierścień kurczliwy powstaje w anafazie, a jego obkurczenie w się z początkiem telofazy prowadzi do powstania bruzdy podziałowej.

27
W procesie cytokinezy poza bruzdą podziałową biorą także udział pęcherzyki i zbiorniki siateczki śródplazmatycznej układające się w płaszczyźnie podziału cytoplazmy i łączą się ze sobą, pomagając w rozdzieleniu cytoplazmy. Prawidłowa kariokineza przy zahamowanej cytokinezie (cytohalazyna B), prowadzi do powstania komórki 2 – jądrowej.
, prowadzi do powstania komórki 2 – jądrowej.")
28
Komórki potomne

29
Regulacja cyklu komórkowego
Każdy narząd dorosłego organizmu ma charakterystyczny wskaźnik mitotyczny. Wskaźnik ten może ulegać wahaniom pod wpływem różnych czynników, ale w warunkach prawidłowych wraca do wartości wyjściowych. Stan, w którym mimo zmieniających się warunków wskaźniki mitotyczne pozostają takie same nosi nazwę homeostazy mitotycznej. Dzięki niej liczba komórek przypadających na określony narząd organizmu dorosłego nie zmienia się. To z kolei decyduje o prawidłowości funkcjonowania narządu. Kiedy liczba komórek układu osiąga wartość charakterystyczną dla organizmu dorosłego, liczba komórek znajdujących się w cyklu ustala się na stałym poziomie.

30
Kinazy cyklu komórkowego
Regulacja cyklu komórkowego odbywa się poprzez uruchomienie kaskadowych reakcji fosforylacji i defosforylacji. Fosforylacja jest katalizowana przez kinazy białkowe, a defosforylacja przez fosfatazy. Substratami kinaz są różne białka jądra i cytoplazmy, w których fosforylacji ulegają najczęściej tyrozyna, seryna i treonina. Aktywacja kinaz zachodzi w dwóch krytycznych przedziałach masowych cyklu komórkowego: pod koniec fazy G2 (co prowadzi do przejścia G1 w S i zapoczątkowania syntezy DNA). Krytyczny przedział masowy wczesnej fazy G1, w którym komórka decyduje się na podział, nazywamy punktem restrykcyjnym.
. Krytyczny przedział masowy wczesnej fazy G1, w którym komórka decyduje się na podział, nazywamy punktem restrykcyjnym.")
31
Regulacja wejścia w mitozę
Przejście późnej fazy G2 w mitozę dokonuje się przez aktywację kinazy fazy M (czynnik aktywujący dojrzewanie MPF – maturation promoting factor). Kinaza fazy M, czyli MPF jest heterodimerem białkowym, składającym się z białka o masie 34 kD i białka o masie 45 kD. Pierwsze to białko p34, drugie to cyklina. Białko p34 jest kinazą katalizującą fosforylację seryny i tyrozyny wielu białek. Poziom makrocząsteczek p34 w komórce, w czasie cyklu komórkowego, jest stały.
. Kinaza fazy M, czyli MPF jest heterodimerem białkowym, składającym się z białka o masie 34 kD i białka o masie 45 kD. Pierwsze to białko p34, drugie to cyklina. Białko p34 jest kinazą katalizującą fosforylację seryny i tyrozyny wielu białek. Poziom makrocząsteczek p34 w komórce, w czasie cyklu komórkowego, jest stały.")
32
Regulacja wejścia w mitozę
Cykliny istnieją w komórkach wielu organizmów jako cykliny A i B, różniące się sekwencją aminokwasów. W czasie cyklu komórkowego, począwszy od wczesnej fazy G1, cykliny są syntetyzowane de novo i dlatego ich poziom w komórkach rośnie w miarę upływu cyklu. Cykliny osiągają maksymalne stężenie w metafazie, po czym ulegają gwałtownemu rozkładowi, prawdopodobnie przez trawienie proteinazami.

33
Regulacja fazy S Odbywa się poprzez: kontrolę przechodzenia komórki z G1 w fazę S, oraz kontrolę kończenia syntezy DNA. Przypuszcza się, że białko p34 może łączyc się w fazie G1 z cyklinami A i B lub innymi białkami dając kompleks kinazowy podobny do kinazy fazy M. Do przejścia S w G2 potrzebne są mechanizmy kończące syntezę DNA i zapobiegające ponownej syntezie już zreplikowanych replikonów; biorą tu udział kinazy p-34, cyklina.

34
Substraty kinazy fazy M
Substratami kinazy fazy M są liczne białka jądrowe i niektóre cytoplazmatyczne. Należą do nich Histon H1, białka blaszki jądrowej, nukleolina, białko SW15 (regulator transkrypcji endonukleazy), antygen T (białko aparatu replikacyjnego DNA u Sv40). Fosforylacja białek substratowych przez kinazę fazy M i defosforylacja przez fosfatazę może powodować kondensację i dekondensację chromatyny, fragmentację i odbudowanie otoczki jądrowej, a także udział w wytwarzaniu wrzeciona podziałowego.
, antygen T (białko aparatu replikacyjnego DNA u Sv40). Fosforylacja białek substratowych przez kinazę fazy M i defosforylacja przez fosfatazę może powodować kondensację i dekondensację chromatyny, fragmentację i odbudowanie otoczki jądrowej, a także udział w wytwarzaniu wrzeciona podziałowego.")
35
Przekazywanie sygnału do wzrostu
Komórki mogą być aktywowane przy pomocy wielu sygnałów wewnętrznych, wśród których należy wymienić czynniki wzrostu, neuromediatory, hormony, bezpośrednie kontakty między komórkami oraz oddziaływanie istoty międzykomórkowej. Sygnały te oddziaływują poprzez receptory powierzchniowe czyli błonowe lub cytoplazmatyczne.

Podobne prezentacje