Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
Przygotowanie wektora do klonowania.
Ćwiczenie 4 Przygotowanie wektora do klonowania. Hydroliza plazmidu pUC19 enzymami HindIII oraz EcoRI. Oczyszczanie DNA wektora po reakcjach enzymatycznych.

2
Hydroliza DNA wektora enzymami restrykcyjnymi HindIII oraz EcoRI
Protokół 4 Hydroliza DNA wektora enzymami restrykcyjnymi HindIII oraz EcoRI Mieszanina reakcyjna (30μl) 1. wektor pUC µl 2. woda dejonizowana 20,5 µl 3. bufor R µl 4. HindIII 1 µl 5. EcoRI 0,5 µl Mieszaninę zwirować (3000obr/min); inkubować 1 godzinę w temp. 37˚C
 1. wektor pUC19 5 µl. 2. woda dejonizowana 20,5 µl. 3. bufor R+ 3 µl. 4. HindIII 1 µl. 5. EcoRI 0,5 µl. Mieszaninę zwirować (3000obr/min); inkubować 1 godzinę w temp. 37˚C.")
3
Protokół 4a Oczyszczanie DNA wektora plazmidowego po trawieniu enzymami - DNA Clean-Up Do probówki zawierającej DNA dodać 150 µl roztworu G i wymieszać przez odwracanie probówki lub worteksowanie. Całość nanieść na minikolumnę do oczyszczania DNA i wirować przy 6000 RPM przez 30s Wyciągnąć kolumnę z probówki, wylać przesącz i ponownie włożyć kolumnę do probówki; dodać do kolumny 600 µl roztworu płuczącego A1 i wirować j.w. Wyciągnąć kolumnę z probówki, wylać przesącz i ponownie włożyć kolumnę do probówki; dodać do kolumny 300 µl roztworu A1 i wirować przy RPM przez 30s. Osuszoną kolumnę umieścić w nowej probówce 1,5 ml i do złoża znajdującego się na dnie kolumny dodać 30 µl buforu TE (lub jałowej wody destylowanej ); Inkubować próbkę 3 min w temperaturze pokojowej, a następnie wirować przy RPM przez 30s.
; Inkubować próbkę 3 min w temperaturze pokojowej, a następnie wirować przy RPM przez 30s.")
4
Wektory i metody wprowadzania wektorów do komórek biorców
klonowany DNA (np. genomowy) musi być na początku procedury klonowania strawiony enzymami restrykcyjnymi. Następnie mieszany jest in vitro z wektorami przeciętymi odpowiednim enzymem, a następnie (po defosforylacji) łączony jest z DNA wektora (ligowany) przy udziale ligazy. W kolejnym etapie musi nastąpić selekcja klonów rekombinowanych. Do tego celu niezbędne jest wstawienie do wektorów tzw. markerów selekcyjnych (genów warunkujących wystąpienie cechy fenotypowej np. gen oporności na tetracyklinę).
 musi być na początku procedury klonowania strawiony enzymami restrykcyjnymi. Następnie mieszany jest in vitro z wektorami przeciętymi odpowiednim enzymem, a następnie (po defosforylacji) łączony jest z DNA wektora (ligowany) przy udziale ligazy. W kolejnym etapie musi nastąpić selekcja klonów rekombinowanych. Do tego celu niezbędne jest wstawienie do wektorów tzw. markerów selekcyjnych (genów warunkujących wystąpienie cechy fenotypowej np. gen oporności na tetracyklinę).")
5
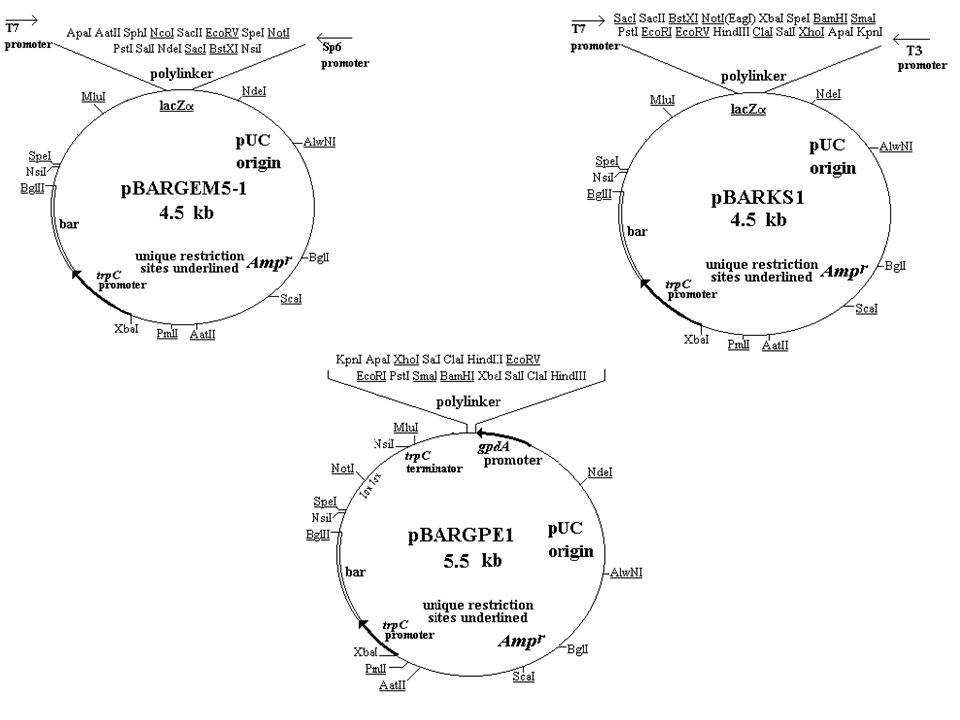
Co to jest wektor? Wektorem służącym do klonowania DNA może być taki element genetyczny, który ma zdolność do namnażania się po wprowadzeniu do komórki biorcy. Wektory powinny ponadto: zawierać geny, tzw. markery selekcyjne, nadające komórkom gospodarza łatwy do testowania fenotyp, np. oporność na antybiotyk, zdolność do rozkładania barwnego substratu zawartego z podłoża itp. być niezdolnym do przeżycia poza komórką gospodarza, namnażać się w niewielu szczepach, to znaczy mieć tzw. mały zakres gospodarza. Rozróżniamy wektory: autonomiczne – replikują one niezależnie (autonomicznie) w komórkach biorcy integracyjne – włączają się (integrują) do DNA gospodarza. W komórkach prokariotycznych używamy najczęściej 3 rodzajów wektorów: Plazmidów, kosmidów, wektorów fagowych
 w komórkach biorcy. integracyjne – włączają się (integrują) do DNA gospodarza. W komórkach prokariotycznych używamy najczęściej 3 rodzajów wektorów: Plazmidów, kosmidów, wektorów fagowych.")
6
WEKTORY PLAZMIDOWE Wektory plazmidowe skonstruowane są z fragmentów naturalnie występujących plazmidów, a ostatnio coraz częściej także z odcinków DNA chromosomowego, fagowego lub syntetycznych oligonukleotydów Klasycznym przykładem wektora plazmidowego jest pBR322, replikujący się w liczbie kopii w komórkach bakterii gramujemnych. Figure pBR322. The map shows the positions of the ampicillin-resistance gene (amp R ), the tetracycline-resistance gene (tet R ), the origin of replication (ori) and the recognition sequences for seven restriction endonucleases (Genomes)
, the tetracycline-resistance gene (tet R ), the origin of replication (ori) and the recognition sequences for seven restriction endonucleases (Genomes)")
7
Konstrukcja nowych wektorów poszła w kierunku:
zmniejszania wielkości wektora (2-10 kb) – małe wymiary plazmidu zwiększają jego pojemność (można klonować wówczas większe fragmenty DNA) oraz wpływają ponadto na zwiększenie częstości transformacji komórek biorcy. Niewielkie plazmidy występują w większej liczbie kopii (nawet /komórkę) włączenia w miejsce klonowania DNA syntetycznego oligonukleotydu, tzw. „polilinkera”, z sekwencjami rozpoznawanymi przez wiele enzymów restrykcyjnych (ang. multiple cloning sites); umożliwia to bezpośrednie wstawienie fragmentów klonowanego DNA, które powstały przez trawienie jednym z wielu lub też dwoma różnymi enzymami. Wycinając następnie tak klonowany DNA innymi ER powodujemy modyfikację jego końców tj. dodanie syntetycznych sekwencji włączenia miejsca klonowania w obrębie genu selekcyjnego, w celu łatwiejszego rozróżnienia komórki z wektorem zrekombinowanym i niezrekombinowanym. Plazmid powinien zawierać wówczas przynajmniej dwa różne markery (geny) selekcyjne. Insercja (wstawienie) obcych sekwencji do jednego z nich inaktywuje go. Brak ekspresji genu selekcyjnego w komórkach transformowanych wektorem, w odpowiednich warunkach selekcyjnych, wskazuje na występowanie w nich wektora zrekombinowanego. dołączenia do wektorów sekwencji ułatwiających selekcję i zwiększających możliwości analizy klonowanych fragmentów
 – małe wymiary plazmidu zwiększają jego pojemność (można klonować wówczas większe fragmenty DNA) oraz wpływają ponadto na zwiększenie częstości transformacji komórek biorcy. Niewielkie plazmidy występują w większej liczbie kopii (nawet /komórkę) włączenia w miejsce klonowania DNA syntetycznego oligonukleotydu, tzw. „polilinkera , z sekwencjami rozpoznawanymi przez wiele enzymów restrykcyjnych (ang. multiple cloning sites); umożliwia to bezpośrednie wstawienie fragmentów klonowanego DNA, które powstały przez trawienie jednym z wielu lub też dwoma różnymi enzymami. Wycinając następnie tak klonowany DNA innymi ER powodujemy modyfikację jego końców tj. dodanie syntetycznych sekwencji. włączenia miejsca klonowania w obrębie genu selekcyjnego, w celu łatwiejszego rozróżnienia komórki z wektorem zrekombinowanym i niezrekombinowanym. Plazmid powinien zawierać wówczas przynajmniej dwa różne markery (geny) selekcyjne. Insercja (wstawienie) obcych sekwencji do jednego z nich inaktywuje go. Brak ekspresji genu selekcyjnego w komórkach transformowanych wektorem, w odpowiednich warunkach selekcyjnych, wskazuje na występowanie w nich wektora zrekombinowanego. dołączenia do wektorów sekwencji ułatwiających selekcję i zwiększających możliwości analizy klonowanych fragmentów.")
10
Figure 4. 18. Recombinant selection with pBR322. See text for details
Figure Recombinant selection with pBR322. See text for details. The inset shows how replica plating is performed.

11
IDENTYFIKACJA KLONÓW ZREKOMBINOWANYCH
wizualna identyfikacja klonów zrekombinowanych możliwa jest po włączeniu do wektora genu kodującego enzym, który rozkłada barwny substrat. Miejsce klonowania obcego DNA znajduje się w obrębie tego genu – przykład pUC19, zawierający sekwencje regulatorowe i N – końcowy fragment genu ß-galaktozydazy (lacZ). Wektory takie wprowadzane są do komórek zawierających zmutowany gen lacZ z delecją w początkowych sekwencjach, które zdolne są do syntezy jedynie C-końcowego fragmentu ß-galoktyzydazy. Obydwa powstałe peptydy mogą asocjować tworząc enzymatycznie aktywne białko (λ-komplementacja). Insercja klonowanego odcinka w obrębie genu lacZ przerywa jego ciągłość i uniemożliwia syntezę peptydu zdolnego do tworzenia aktywnej cząsteczki ß-galaktozydazy. Komórki produkujące aktywny enzym hydrolizują substrat zawarty w podłożu, nadając barwę koloniom na agarze. Zrekombinowane komórki nie posiadają takiej zdolności (kolonie są białe). selekcja zrekombinowanych klonów może zachodzić bezpośrednio poprzez zahamowanie namnażania się komórek posiadających niezrekombinowany plazmid. Do tak działających wektorów włączony jest gen letalny dla komórki gospodarza. Insercja klonowanego DNA w obrębie tego genu inaktywuje ekspresję, umożliwiając namnażanie się rekombinantów
. Wektory takie wprowadzane są do komórek zawierających zmutowany gen lacZ z delecją w początkowych sekwencjach, które zdolne są do syntezy jedynie C-końcowego fragmentu ß-galoktyzydazy. Obydwa powstałe peptydy mogą asocjować tworząc enzymatycznie aktywne białko (λ-komplementacja). Insercja klonowanego odcinka w obrębie genu lacZ przerywa jego ciągłość i uniemożliwia syntezę peptydu zdolnego do tworzenia aktywnej cząsteczki ß-galaktozydazy. Komórki produkujące aktywny enzym hydrolizują substrat zawarty w podłożu, nadając barwę koloniom na agarze. Zrekombinowane komórki nie posiadają takiej zdolności (kolonie są białe). selekcja zrekombinowanych klonów może zachodzić bezpośrednio poprzez zahamowanie namnażania się komórek posiadających niezrekombinowany plazmid. Do tak działających wektorów włączony jest gen letalny dla komórki gospodarza. Insercja klonowanego DNA w obrębie tego genu inaktywuje ekspresję, umożliwiając namnażanie się rekombinantów.")
12
EKSPRESJA otrzymanie dużych ilości mRNA syntetyzowanego na matrycy klonowanego DNA możliwe jest, gdy wektor zaopatrzony jest w silny promotor rozpoznawany przez polimerazę RNA gospodarza (wektory ekspresyjne). zawierają one także często sekwencje regulujące transkrypcję. Klonowanie genów eukariotycznych w komórkach prokariotycznych wymaga wprowadzenia genów eukariotycznych pod kontrolę prokariotycznych sekwencji regulujących – operatora i promotora. do konstrukcji wektorów ekspresyjnych stosuje się m.in. promotory: pL faga lambda, lac E.coli, trp E.coli i bla (promotor genu ß-laktamazy). Promotor pL faga lambda jest zwykle przynajmniej kilkakrotnie bardziej efektywny niż promotor lac. Kontrolowany jest on przez wrażliwy na wysoką temperaturę represor CI. W podwyższonej temperaturze następuje inaktywacja represora, co powoduje wzrost ekspresji genu. Geny z promotorem trp indukowane są analogiem tryptofanu. Ich ekspresja może prowadzić do syntezy nawet 20% białka komórkowego;
. zawierają one także często sekwencje regulujące transkrypcję. Klonowanie genów eukariotycznych w komórkach prokariotycznych wymaga wprowadzenia genów eukariotycznych pod kontrolę prokariotycznych sekwencji regulujących – operatora i promotora. do konstrukcji wektorów ekspresyjnych stosuje się m.in. promotory: pL faga lambda, lac E.coli, trp E.coli i bla (promotor genu ß-laktamazy). Promotor pL faga lambda jest zwykle przynajmniej kilkakrotnie bardziej efektywny niż promotor lac. Kontrolowany jest on przez wrażliwy na wysoką temperaturę represor CI. W podwyższonej temperaturze następuje inaktywacja represora, co powoduje wzrost ekspresji genu. Geny z promotorem trp indukowane są analogiem tryptofanu. Ich ekspresja może prowadzić do syntezy nawet 20% białka komórkowego;")
13
jednoniciowe kopie DNA plazmidu można otrzymać przez włączenie do wektora fragmentu ori, tj. początku replikacji pochodzącego z jednoniciowego bakteriofaga np. M13. Jednoniciowe matryce stosowane są do sekwencjonowania DNA. Przkład plazmidu – Bluescript M13 (2,96 kb). wektory umożliwiające transkrypcję klonowanego DNA in vitro zawierają promotory bakteriofagów, np. T3, T7 lub SP6, umieszczone przed miejscem wstawienia DNA. Po linearyzacji plazmidu inkubuje się go z odpowiednią dla danego promotora polimerazą RNA zależną od DNA i rybonukleotydami. Tak otrzymane próbki można następnie zastosować do translacji w systemie bezkomórkowym. Przykład – pGEM-3 (2867 bp).
.")
14
KOSMIDY Kosmidy są zmodyfikowanymi plazmidami, zawierającymi oprócz fragmentu ori, genu selekcyjnego i miejsca do klonowania także sekwencje cos faga λ. W normalnym cyklu życiowym faga λ, DNA jest syntetyzowany w postaci długich cząsteczek, konkatamerów, w których genom bakteriofaga jest powtórzony wielokrotnie. Podczas pakowania do główek cząsteczki te są cięte w miejscach cos przy udziale fagowego białka A, na odcinki DNA odpowiadające długości DNA bakteriofaga. Ponieważ kosmidy mają około 5 kb, długość klonowanych w nich fragmentów DNA musi wynosić około kb. Cząsteczki kosmidów przecinane są ER i łączone z fragmentami obcego (klonowanego) DNA. Powstałe długie odcinki DNA, w których klonowany DNA przedzielony jest cząsteczką kosmidu, ulegają fragmentacji w miejscach cos i są upakowywane w kapsydach. Po infekcji szczepu E.coli zrekombinowany, liniowy DNA wstrzykiwany jest do komórek, gdzie podlega cyrkularyzacji, tj. łączą się jego lepkie końce cos. Koliste cząsteczki kosmodu replikują się i nadają nowe cechy komórkom gospodarza pozwalające na ich selekcję.
 DNA. Powstałe długie odcinki DNA, w których klonowany DNA przedzielony jest cząsteczką kosmidu, ulegają fragmentacji w miejscach cos i są upakowywane w kapsydach. Po infekcji szczepu E.coli zrekombinowany, liniowy DNA wstrzykiwany jest do komórek, gdzie podlega cyrkularyzacji, tj. łączą się jego lepkie końce cos. Koliste cząsteczki kosmodu replikują się i nadają nowe cechy komórkom gospodarza pozwalające na ich selekcję.")
15
KOSMIDY Kosmidy służą do klonowania stosunkowo długich cząsteczek DNA (33-47 kb), zbyt dużych do przenoszenia w wektorach pochodnych faga λ lub wektorach plazmidowych. Są one szczególnie przydatne do badania wielu genów eukariotycznych, zawierających liczne, długie introny, oraz innych sekwencji genomowego DNA.
, zbyt dużych do przenoszenia w wektorach pochodnych faga λ lub wektorach plazmidowych. Są one szczególnie przydatne do badania wielu genów eukariotycznych, zawierających liczne, długie introny, oraz innych sekwencji genomowego DNA.")
16
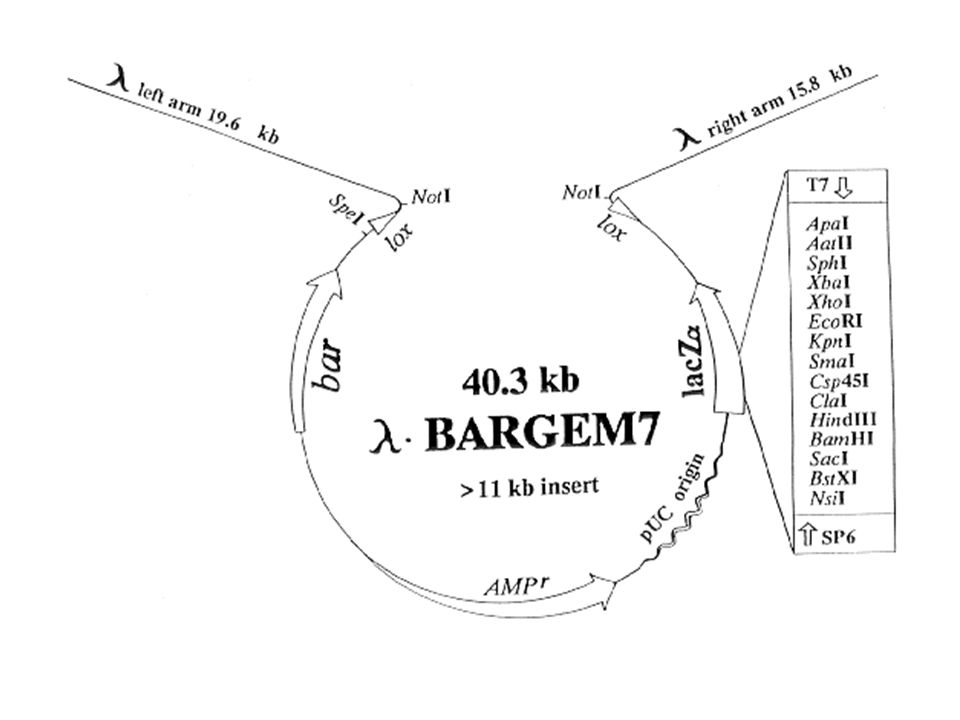
WEKTORY FAGOWE Dwa typy fagów E. Coli znalazły ogromne zastosowanie jako prokariotyczne wektory do klonowania, są to: fag λ jednoniciowy fag M13 Fag λ zawiera liniowy, 2-niciowy DNA o długości par zasad z jednoniciowymi lepkimi końcami cos, długości 12 nukleotydów. Liczne genetyczne zmiany, wprowadzone w DNA faga w wielu laboratoriach, doprowadziły do otrzymania znacznej liczby pochodnych faga λ (>100), używanych jako wektory do klonowania. Fag λ typu dzikiego nie może być stosowany jako wektor. Jednym z powodów tego jest brak unikalnych miejsc rozpoznawanych przez ER. Drugą niedogodnością cząsteczki DNA faga λ typu dzikiego jako wektora jest bardzo ograniczona wielkość fragmentu obcego DNA, który może być wklonowany (nie więcej niż 10% genomu dzikiego λ). Koniecznym było zatem skonstruowanie fagów delecyjnych.
, używanych jako wektory do klonowania. Fag λ typu dzikiego nie może być stosowany jako wektor. Jednym z powodów tego jest brak unikalnych miejsc rozpoznawanych przez ER. Drugą niedogodnością cząsteczki DNA faga λ typu dzikiego jako wektora jest bardzo ograniczona wielkość fragmentu obcego DNA, który może być wklonowany (nie więcej niż 10% genomu dzikiego λ). Koniecznym było zatem skonstruowanie fagów delecyjnych.")
17
Ponieważ część genomu faga λ jest nieistotna dla cyklu litycznego, skontruowano takie mutanty delecyjne, które zwiększały możliwość sklonowania większych fragmentów obcego DNA. Wśród nich były również takie, które posiadały bardzo rozległe delecje (nawet do 25% genomu faga). Taki DNA (delecyjny) nie może być pakowany do główek fagowych, co staje się zaletą w procedurze klonowania, gdyż tylko DNA z wklonowanym obcym DNA może się pakować (dobra selekcja fagów zrekombinowanych). Około 60% genomu faga niezbędne jest do jego litycznego wzrostu. Zawiera ona geny syntezy główki i ogonka faga oraz region replikacji, geny lityczne i sekwencje cos. Środkowa część genomu faga , około 1/3 całości cząsteczki DNA może być zastąpiona przez fragment obcego DNA (klonowanego) lub wstawione geny selekcyjne. Zrekombinowane fagi mogą być selekcjonowane ma wiele sposobów: selekcja wg wielkości, metabolizm laktozy lub biotyny morfologia łysinek fagowych, analiza restrykcyjna
. Taki DNA (delecyjny) nie może być pakowany do główek fagowych, co staje się zaletą w procedurze klonowania, gdyż tylko DNA z wklonowanym obcym DNA może się pakować (dobra selekcja fagów zrekombinowanych). Około 60% genomu faga niezbędne jest do jego litycznego wzrostu. Zawiera ona geny syntezy główki i ogonka faga oraz region replikacji, geny lityczne i sekwencje cos. Środkowa część genomu faga , około 1/3 całości cząsteczki DNA może być zastąpiona przez fragment obcego DNA (klonowanego) lub wstawione geny selekcyjne. Zrekombinowane fagi mogą być selekcjonowane ma wiele sposobów: selekcja wg wielkości, metabolizm laktozy lub biotyny. morfologia łysinek fagowych, analiza restrykcyjna.")
18
Rycina. LambdaGem-11 Genomic Cloning

19
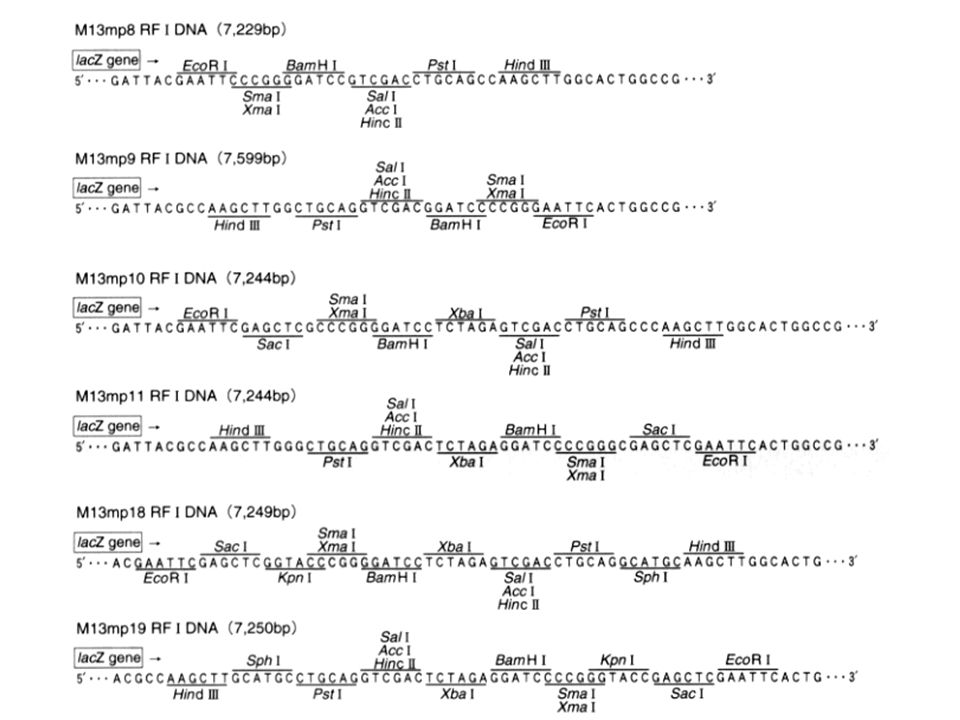
Fag M13 zbudowany jest z jednoniciowego DNA (6407 nukleotydów), bardzo mały w stosunku do genomu faga λ, występuje w komórce w liczbie kopii. Po wniknięciu DNA faga M13 do komórki E.coli dosyntetyzowana jest druga, komplementarna nić DNA, a następnie zachodzi replikacja formy dwuniciowej. W końcowej fazie infekcji powstają jednoniciowe cząsteczki, które łączą się z osłonkami białkowymi i wydostają z komórki nie powodując jej lizy. Osłonka faga M13 nie limituje wielkości upakowanych cząsteczek, jednak wstawki większe niż 1 kb wykazują niestabilność w tego typu cząsteczkach. Fag M13 ma 10 genów, z których żaden nie może zostać usunięty lub unieczynniony bez wpływu na żywotność faga. Po długich badaniach stwierdzono, że w jedno z 10 miejsc rozpoznawanych przez ER BsuI można wstawić fragment obcego DNA nie powodując zaburzenia funkcji życiowych. W miejsce to wstawiono fragment operonu laktozowego E.coli (miejsce dla ER EcoRI). W ten sposób skonstruowano szereg pochodnych faga M13, wykorzystywanych do klonowania – tzw. wektory M13mp. Wektory te posiadają promotor i N-końcowy fragment genu ß-galaktozydazy z E.coli, z wbudowanym polilinkerem (MCS). Umożliwia to bezpośrednią selekcję klonów zrekombinowanych. Wektory powstałe na bazie faga M13 umożliwiają bezpośrednie sekwencjonowanie nukleotydów klonowanego odcinka DNA.
. W ten sposób skonstruowano szereg pochodnych faga M13, wykorzystywanych do klonowania – tzw. wektory M13mp. Wektory te posiadają promotor i N-końcowy fragment genu ß-galaktozydazy z E.coli, z wbudowanym polilinkerem (MCS). Umożliwia to bezpośrednią selekcję klonów zrekombinowanych. Wektory powstałe na bazie faga M13 umożliwiają bezpośrednie sekwencjonowanie nukleotydów klonowanego odcinka DNA.")
21
WEKTORY WAHADŁOWE Wektory wahadłowe replikują się w różnych gatunkach mikroorganizmów prokariotycznych jak i eukariotycznych komórkach gospodarzy. Typowy wektor wahadłowy skonstruowany jest z części prokariotycznej i eukariotycznej. Część prokariotyczna wektora zawiera: sekwencję ori (start replikacji) marker selekcyjny Część eukariotyczna wektora zawiera: sekwencję ori rozpoznawaną przez polimerazę DNA gospodarza eukariotycznego sekwencje umożliwiające ekspresję genu – promotor, „polilinker”, sekwencja intronowa, terminator. Najczęściej stosowanymi wektorami wahadłowymi są wektory drożdżowe. Większość z nich zawiera sekwencje pochodzące z plazmidów bakteryjnych (pBR 322 i jego pochodnych) oraz sekwencje drożdżowe. Wektory te mogą być użyte jako wahadłowe, tzn. zrekombinowany gen można za ich pośrednictwem wprowadzić zarówno do komórek E.coli jak i komórek drożdży
 marker selekcyjny. Część eukariotyczna wektora zawiera: sekwencję ori rozpoznawaną przez polimerazę DNA gospodarza eukariotycznego. sekwencje umożliwiające ekspresję genu – promotor, „polilinker , sekwencja intronowa, terminator. Najczęściej stosowanymi wektorami wahadłowymi są wektory drożdżowe. Większość z nich zawiera sekwencje pochodzące z plazmidów bakteryjnych (pBR 322 i jego pochodnych) oraz sekwencje drożdżowe. Wektory te mogą być użyte jako wahadłowe, tzn. zrekombinowany gen można za ich pośrednictwem wprowadzić zarówno do komórek E.coli jak i komórek drożdży.")
Podobne prezentacje