Pobierz prezentację
This is a modal window.
1
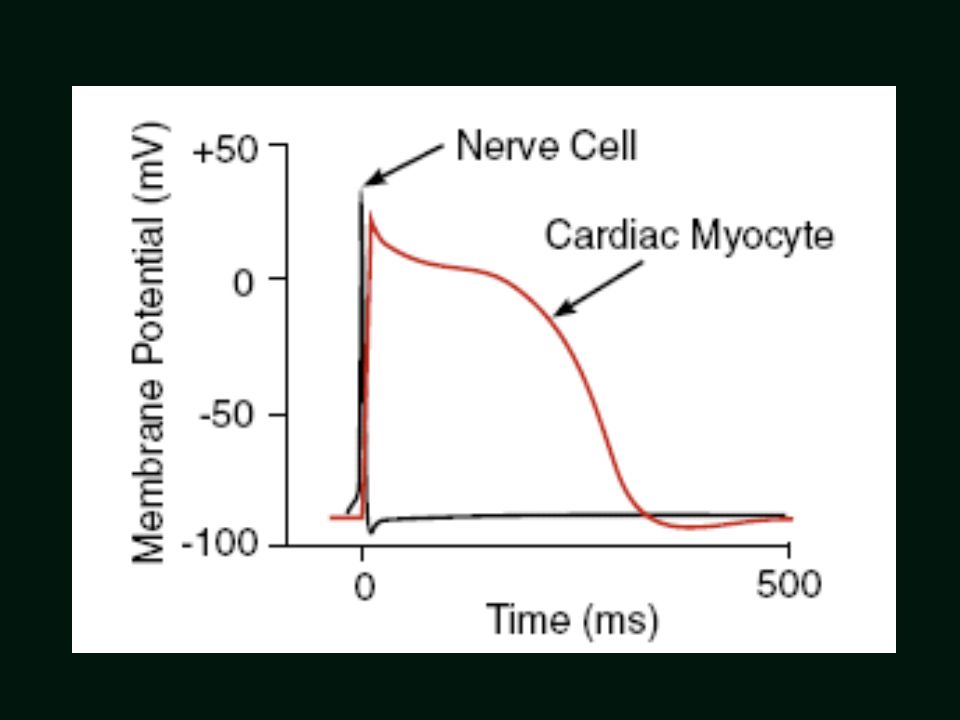
Elektrofizjogiczne uwarunkowania czynności serca
Tomasz H. Wierzba Elektrofizjogiczne uwarunkowania czynności serca 23-II-2012 Hard Heart 1. Dee Unglaub Silverthorn – Human Physiology; Pearson Int. 2007; 2. Ole H. Petersen: Human Physiology, Lecture Notes; Blackwell 2007; 3. A.C. Gyuton & J.E. Hall: Textbook of Medical Physiology; Elsevier, 2006; 4. W.F. Boron & E.L. Boulpaep: Medical Physiology, Elsevier, Saunders, 2005; 5. L.S.Constanzo Physiology, Lippincott Willains and Wilkins, 2007; 6. Sabyasachi Sircar: Principles of Medical Physiology, Thieme, 2008; 7. S.J. Konturek (red.) Fizjologia człowieka, Urban & Partner, 2007; 8. J. Górski (red.) Fizjologiczne podstawy wysiłku fizycznego, PZWL 2006; 9. W.Z. Traczyk, A. Trzebski (red.): Fizjologia człowieka z elementami fizjologii stosowanej i klinicznej, PZWL, 2001
 Fizjologia człowieka, Urban & Partner, 2007; 8. J. Górski (red.) Fizjologiczne podstawy wysiłku fizycznego, PZWL 2006; 9. W.Z. Traczyk, A. Trzebski (red.): Fizjologia człowieka z elementami fizjologii stosowanej i klinicznej, PZWL,")
2
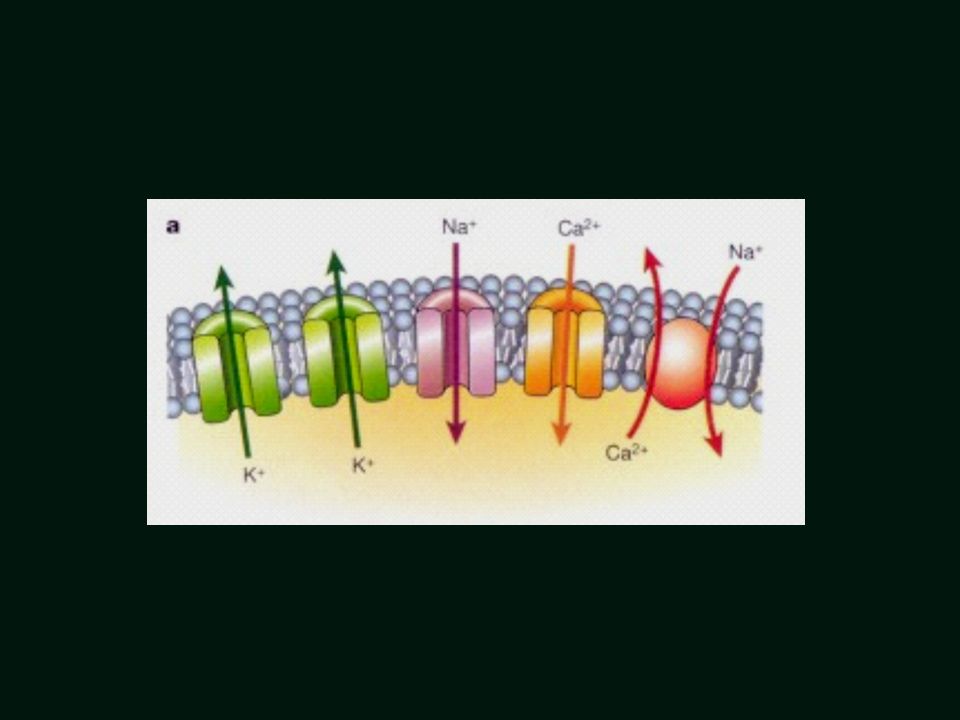
Pompa sodowo-potasowa – umożliwia ruch jonów w poprzek błony wbrew gradientowi stężeń

3
Geneza potencjału spoczynkowego

6
Potencjał równowagi dla danego jonu →

7
Potencjał równowagi dla danego jonu
Równanie Nernsta-Goldmana (opisane także niezależnie przez: Hodgkina i Katza) Przeciętnie: Na mV K mV Cl mV
 Przeciętnie: Na mV. K mV. Cl mV.")
8
Potencjał błonowy [mV] - 55 - 70 Czas
![Potencjał błonowy [mV] Czas](http://slideplayer.pl/slide/440066/1/images/8/Potencja%C5%82+b%C5%82onowy+%5BmV%5D+Czas.jpg "Potencjał błonowy [mV] Czas")
9
Potencjał czynnościowy – zasada „wszystko albo nic” (0 lub 1)
Potencjał błonowy [mV] - 55 - 70 Czas

10
Potencjał błonowy [mV] - 55 - 70 Czas
![Potencjał błonowy [mV] Czas](http://slideplayer.pl/slide/440066/1/images/10/Potencja%C5%82+b%C5%82onowy+%5BmV%5D+Czas.jpg "Potencjał błonowy [mV] Czas")
14
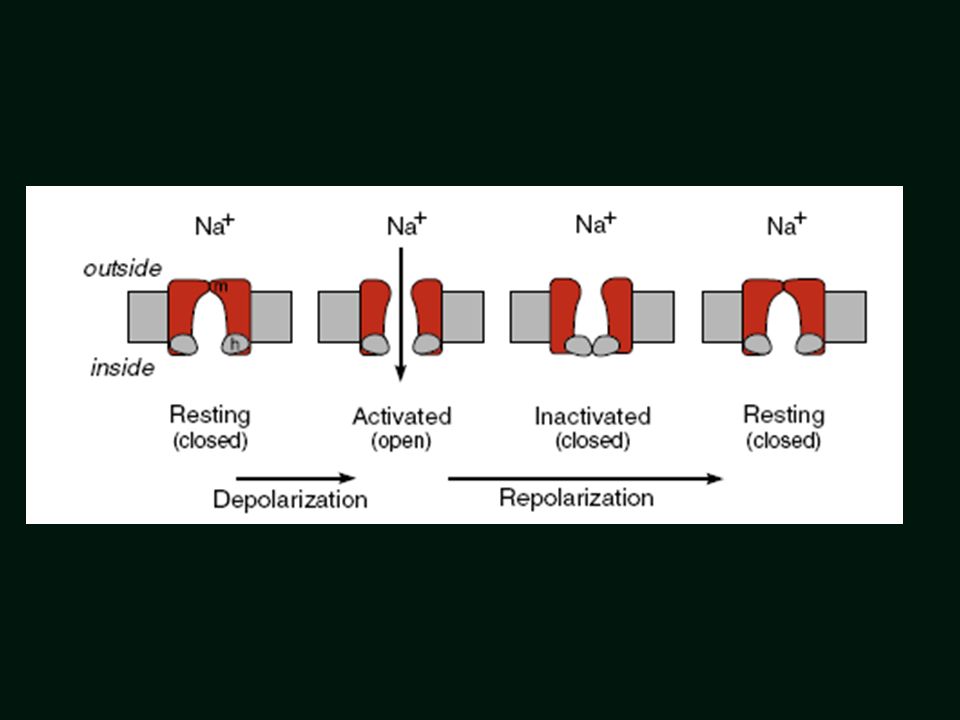
Zmiana konfiguracji kanałów Na+
= Outward K+ Current Kanały Na+ zaktywowane Zmiana konfiguracji kanałów Na+

15
iCa++ iNa+

16
Fizjologiczna aktywacja mięśnia sercowego
Neuromuscular Junction Action Potential Sarcolemma AChRs Ca2+ Sarcoplasmic Reticulum RyR Transverse Tubule DHPR Ca2+ Troponin Actin Tropomyosin Myosin 16 16 16

17
17

18
DHPR – receptory dihydropirydynowe
DHPR tworzą kanały wapniowe należące do typu, które są kanałami potencjało-zależnymi, tzn. otwierają się przy przekroczeniu poziomu depolaryzacji progowej Aktywator – depolaryzacja Inhibitor - dihydropirydyna W komórkach mięśnia sercowego DHPR nie są mechanicznie sprzężone z RYR. Analogicznie do mięśni szkieletowych, aktywacja DHPR powoduje napływ Ca2+ do komórki, w obrębie triady. Lokalny wzrost Ca2+ powoduje otwarcie RYR i wypływ Ca2+ z SR. Ten mechanizm sprzężenia zwrotnego dodatniego z udziałem Ca2+ jest określany mianem CIRC (Calcium Induced Calcium Release)
")
20
Otwarcie kanału Na+ w fazie 0
“szybki kanał sodowy - iNa+

21
Faza 1 Dokoczenie inaktywacji kanału of Na+
Przejściowy zewnątrzkomórkowy prąd K+

22
Udział jonów Na+ i K+ w Fazie 0 i Fazie 1
Na+ current ends prąd Na+ Outward K+ current

23
Faza 2 (Plateau) Wolny kanał wapniowy (kanał Ca++ typu L)
równoważony przez : Odśrodkowy prąd K+

24
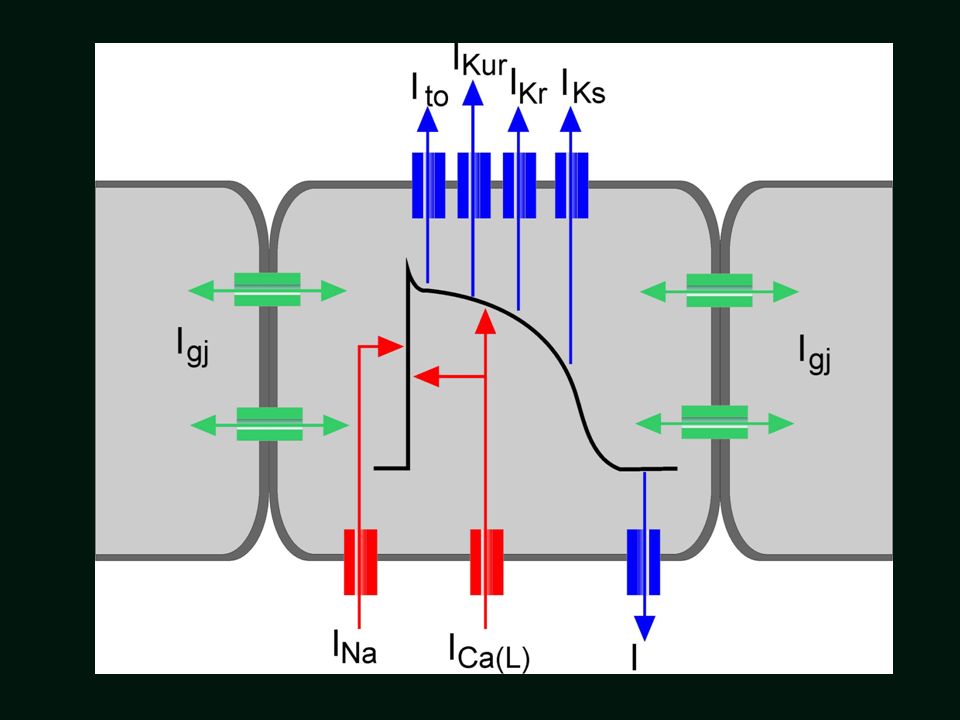
Podstawowe rodzaje prądów potencjału czynnościowego komórek roboczych mięśnia sercowego
Faza 1 (Ito K kanał) -80 mV +60 mV Faza 2 (ICa, IKs, IKr) Faza 3 (IKs, IKr) Faza 0 (INa) Faza 4 (IK1) Faza 4 – potencjał spoczynkowy 24
 -80 mV. +60 mV. Faza 2 (ICa, IKs, IKr) Faza 3 (IKs, IKr) Faza 0. (INa) Faza 4 (IK1) Faza 4 – potencjał spoczynkowy. 24.")
25
Currents that underlie the Cardiac Action Potential (AP)
Phase 1 (KCND) -80 mV +60 mV Faza 2 (CACNA1C, IKs, IKr) Faza 3 (KCNQ1+KCNE1) , KCNH2(HERG)+ KCNE2?) Faza 0 (SCN5A) Faza 4 (KCNJ1) Phase 4 (diastole) 25
 -80 mV. +60 mV. Faza 2 (CACNA1C, IKs, IKr) Faza 3 (KCNQ1+KCNE1) , KCNH2(HERG)+ KCNE2 ) Faza 0. (SCN5A) Faza 4 (KCNJ1) Phase 4 (diastole) 25.")
26
Contribution of Ion channels that underlie the cardiac AP
Prądy depolaryzujące Prądy hiperpolaryzujące lub repolaryzujące

27
Currents in the heart 27

28
Purpose of currents 28

29
Fizjologiczna aktywacja mięśnia sercowego
Neuromuscular Junction Action Potential Sarcolemma AChRs Ca2+ Sarcoplasmic Reticulum RyR Transverse Tubule DHPR Ca2+ Troponin Actin Tropomyosin Myosin 29 29

31
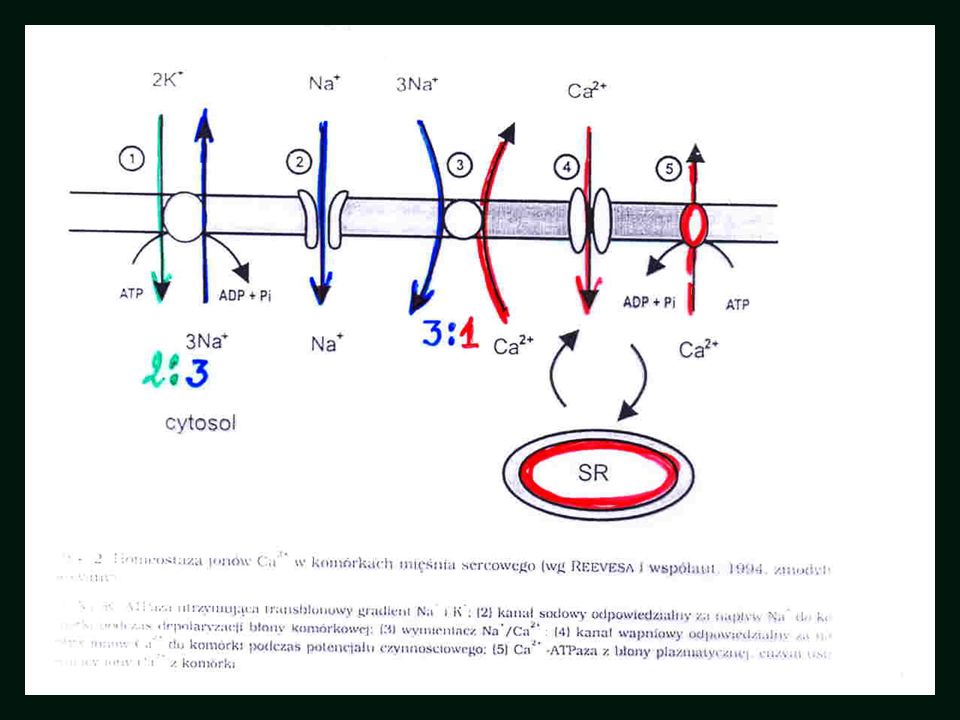
ATP-aza NCX K+ Na+ Na+ Na+ Ca++

32
Potencjał równowagi dla danego jonu
Równanie Nernsta-Goldmana (opisane także niezależnie przez: Hodgkina i Katza)
")
33
Basic Electrophysiology Excitation- Contraction Coupling Regulation of Intracellular Calcium
Cardiac excitation–contraction (E-C) coupling refers to the process whereby myocyte excitation (in the form of membrane depolarization) is used as the stimulus for a coordinated movement of calcium around the cell to bring about contraction. It uses an efficient amplification system, which uses both intracellular and sarcolemmal (membrane) bound components. Cardiac E-C coupling plays a vital role in maintaining the normal inotropic (contractile state) and electrical properties of the heart. It is not surprising that when this process is defective both heart failure and cardiac arrhythmias can result. 1) When the myocyte depolarises, extracellular Ca2+ enters the cell through the L type voltage dependent (called dihydropyridine sensitive because of what causes the membrane channel to open) calcium channel, a phenomenon represented by the phase 2 plateau of the cardiac action potential. 2) Additional calcium is required and this is obtained from a pool of stored calcium within the sarcoplasmic reticulum (SR) of the cell (see figure 1). 3) The initial inward movement of Ca2+ acts as an amplification signal for the release of this storage pool of SR calcium. 4) This process, known as “calcium induced calcium release”, occurs through an SR membrane ion channel known as the cardiac ryanodine receptor (RyR2, the drug that make the receptor change its comformation allowing calcium to be released). 5) Individual populations of RyR2 localise in areas of the SR membrane, adjacent to L type voltage dependent channels within the T tubules of the sarcolemma. Influx of calcium through this latter channel activates its associated local population of RyR2 channels, causing a synchronised release of calcium known as a Ca2+ spark. The synchronised release of multiple Ca2+ sparks throughout the cell following depolarisation creates a global intracellular Ca2+ transient of sufficient magnitude to bring about contraction (slide 14). 6) Conversely, myocyte relaxation results from closure of RyR2 and the rapid removal of cytosolic calcium, either by reuptake into the SR through the sarcoplasmic Ca2+/ATPase pump (SERCA), or by its efflux out of the cell through the sarcolemmal Na+/Ca2+ exchanger (NCX). 7) These two processes balance the earlier cellular influx and SR release of calcium so that there is no net gain or loss of cellular calcium (referred to as homeostatis, what ever goes in must come out) with each contraction–relaxation cycle.
 coupling refers to the process whereby myocyte excitation (in the form of membrane depolarization) is used as the stimulus for a coordinated movement of calcium around the cell to bring about contraction. It uses an efficient amplification system, which uses both intracellular and sarcolemmal (membrane) bound components. Cardiac E-C coupling plays a vital role in maintaining the normal inotropic (contractile state) and electrical properties of the heart. It is not surprising that when this process is defective both heart failure and cardiac arrhythmias can result. 1) When the myocyte depolarises, extracellular Ca2+ enters the cell through the L type voltage dependent (called dihydropyridine sensitive because of what causes the membrane channel to open) calcium channel, a phenomenon represented by the phase 2 plateau of the cardiac action potential. 2) Additional calcium is required and this is obtained from a pool of stored calcium within the sarcoplasmic reticulum (SR) of the cell (see figure 1). 3) The initial inward movement of Ca2+ acts as an amplification signal for the release of this storage pool of SR calcium. 4) This process, known as calcium induced calcium release , occurs through an SR membrane ion channel known as the cardiac ryanodine receptor (RyR2, the drug that make the receptor change its comformation allowing calcium to be released). 5) Individual populations of RyR2 localise in areas of the SR membrane, adjacent to L type voltage dependent channels within the T tubules of the sarcolemma. Influx of calcium through this latter channel activates its associated local population of RyR2 channels, causing a synchronised release of calcium known as a Ca2+ spark. The synchronised release of multiple Ca2+ sparks throughout the cell following depolarisation creates a global intracellular Ca2+ transient of sufficient magnitude to bring about contraction (slide 14). 6) Conversely, myocyte relaxation results from closure of RyR2 and the rapid removal of cytosolic calcium, either by reuptake into the SR through the sarcoplasmic Ca2+/ATPase pump (SERCA), or by its efflux out of the cell through the sarcolemmal Na+/Ca2+ exchanger (NCX). 7) These two processes balance the earlier cellular influx and SR release of calcium so that there is no net gain or loss of cellular calcium (referred to as homeostatis, what ever goes in must come out) with each contraction–relaxation cycle.")
34
NCX – wymiennik Na+/Ca++
Na+ In Cell ECF Nadmiar Ca++ wskutek CIRC Nadmiar Ca++ wskutek oddysocjowania Ca++ od kompleksu aktyna-miozyna Ca++ na zewn.

35
Kanały wapniowe, wymienniki jonowe

36
Heterogenność sercowych potencjałów czynnościowych

38
Ik+ - przykład prądu jonowego aktywowanego depolaryzacją
Zestawienie: potencjał czynnościowy komórek rozrusznika I-rzędowego i mięśnia roboczego komór Szybki prąd sodowy i wolny prąd wapniowy Nie działa szybki prąd sodowy Ik+ - przykład prądu jonowego aktywowanego depolaryzacją If – (f –funny) – dośrodkowy prąd głównie Na+ z dodatkiem (20%) K+ - prąd aktywowany przez hiperpolaryzację.
 – dośrodkowy prąd głównie Na+ z dodatkiem (20%) K+ - prąd aktywowany przez hiperpolaryzację.")
39
Ik+ - przykład prądu jonowego aktywowanego depolaryzacją
If – (f –funny) – dośrodkowy prąd głównie Na+ z dodatkiem (20%) K+ - prąd aktywowany przez hiperpolaryzację. Mechanizmy prepotencjału – powolnej (samoistnej) depolaryzacji spoczynkowej komórek rozrusznikowych Punkt odniesienia: potencjał równowagi dla danego jonu 1. Zamknięcie kanałów potasowych (w fazie 0 i 1 kanały K, które zwykle są otwarte powoli zamykają się – wpływ depolaryzacji; Zatem: mniej jonów K+ opuszcza komórkę = mniej kationów opuszcza komórkę → tendencja do zmniejszenia potencjału (depolaryzacji) komórki (przy równocześnie aktywnych innych prądach tła) 2. If – prąd aktywowany hiperpolaryzacją pod koniec fazy 3: dokomórkowy napływ jonów Na+ i (konwekcyjnie) K+ Skutek: depolaryzacja. Przede wszystkim włókna Purkynjego, ale także komórki rozrusznikowe węzła zatokowego i p-k. 3. Odkomórkowy prąd chlorkowy – występuje w niektórych komórkach, ale wydaje się mieć znaczenie marginalne 4. Dokomórkowy prąd wapniowy – teoretycznie może wywoływać depolaryzację, ale nie stwierdzono, żeby otwarcie błonowych kanałów wapniowych w znaczącym stopniu uczestniczyło w powolnej spoczynkowej depolaryzacji
 – dośrodkowy prąd głównie Na+ z dodatkiem (20%) K+ - prąd aktywowany przez hiperpolaryzację. Mechanizmy prepotencjału – powolnej (samoistnej) depolaryzacji spoczynkowej komórek rozrusznikowych. Punkt odniesienia: potencjał równowagi dla danego jonu. 1. Zamknięcie kanałów potasowych (w fazie 0 i 1 kanały K, które zwykle są otwarte powoli zamykają się – wpływ depolaryzacji; Zatem: mniej jonów K+ opuszcza komórkę = mniej kationów opuszcza komórkę → tendencja do zmniejszenia potencjału (depolaryzacji) komórki (przy równocześnie aktywnych innych prądach tła) 2. If – prąd aktywowany hiperpolaryzacją pod koniec fazy 3: dokomórkowy napływ jonów Na+ i (konwekcyjnie) K+ Skutek: depolaryzacja. Przede wszystkim włókna Purkynjego, ale także komórki rozrusznikowe węzła zatokowego i p-k. 3. Odkomórkowy prąd chlorkowy – występuje w niektórych komórkach, ale wydaje się mieć znaczenie marginalne. 4. Dokomórkowy prąd wapniowy – teoretycznie może wywoływać depolaryzację, ale nie stwierdzono, żeby otwarcie błonowych kanałów wapniowych w znaczącym stopniu uczestniczyło w powolnej spoczynkowej depolaryzacji.")
40
Potencjał czynnościowy komórek rozrusznikowych
Faza 0 Faza 2 b krótka, trudna do wyróżnienia Faza 3 Faza 4 Brak Fazy 1 2 3 4

41
Prądy jonowe - potencjału czynnościowego komórek węzłów SA i AV
Depolarization głownie przez napływ Ca++

42
napływ K+ powoduje repolaryzację
Kationy z sąsiadujących komórek Ca++ channel recovery Relative Refractory Period Ca++ channels activated

43
Okresy refrakcji bezwzględnej (ERP) i względnej (RRP)
 i względnej (RRP)")
44
SERCE - POTENCJAŁY CZYNNOŚCIOWE
Szybko narastający potencjał czynnościowy Wolno narastający potencjał czynnościowy -100 -50 Komórkowy potencjał czynnościowy (mV) Względna przepuszczalność błony komórkowej 10,0 1,0 0,1 -80 -100 -50 Komórkowy potencjał czynnościowy (mV) Względna przepuszczalność błony komórkowej 10,0 1,0 0,1 -80 Faza 1 Faza 2 Faza 0 Faza 3 Faza 4 Faza 4 K+ K+ Na+ Na+ Ca2+ Ca2+ 0,15 0,30 0,15 0,30 Czas (sek) Czas (sek)
 Względna przepuszczalność błony komórkowej. 10,0. 1,0. 0, Komórkowy potencjał czynnościowy (mV) Względna przepuszczalność błony komórkowej. 10,0. 1,0. 0, Faza 1. Faza 2. Faza 0. Faza 3. Faza 4. Faza 4. K+ K+ Na+ Na+ Ca2+ Ca2+ 0,15. 0,30. 0,15. 0,30. Czas (sek) Czas (sek)")
45
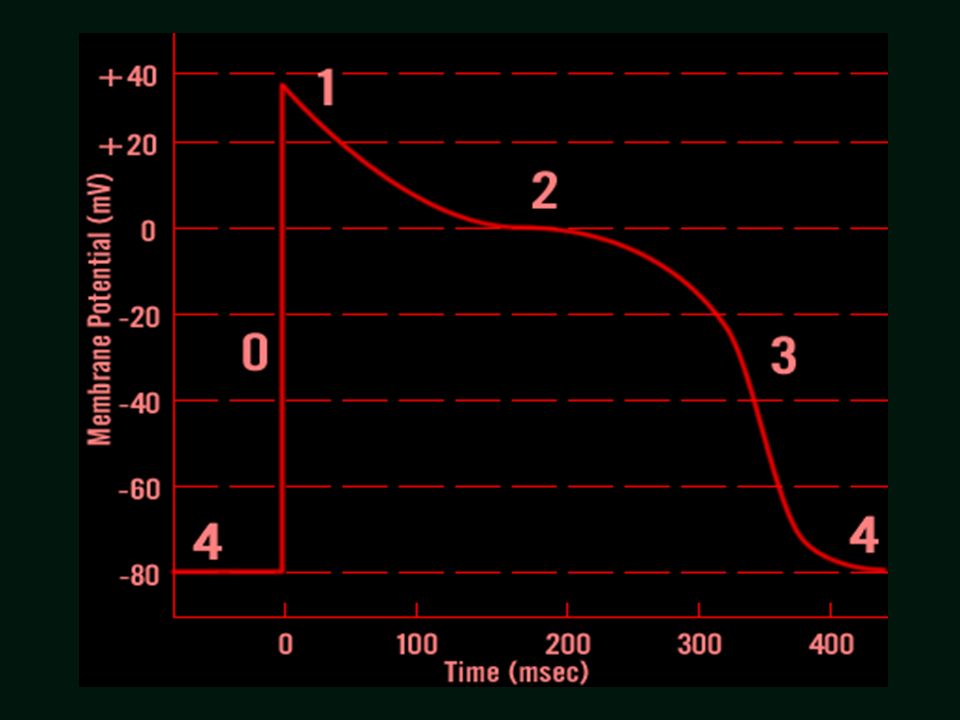
mV +30 1 2 3 4 -90 100% SERCE - POTENCJAŁY CZYNNOŚCIOWE
Potencjał czynnościowy Skurcz mięśnia +30 Faza 0 – depolaryzacja Faza 1 – repolaryzacja wstępna Faza 2 – plateau potencjału Faza 3 – końcowa repolaryzacja Faza 4 – wyjściowy potencjał spoczynkowy 1 2 3 4 -90 100% ORB – okres refrakcji bezwzględnej ORW – okres refrakcji względnej ORC– okres refrakcji czynnościowej czas ORB ORW ORC

46
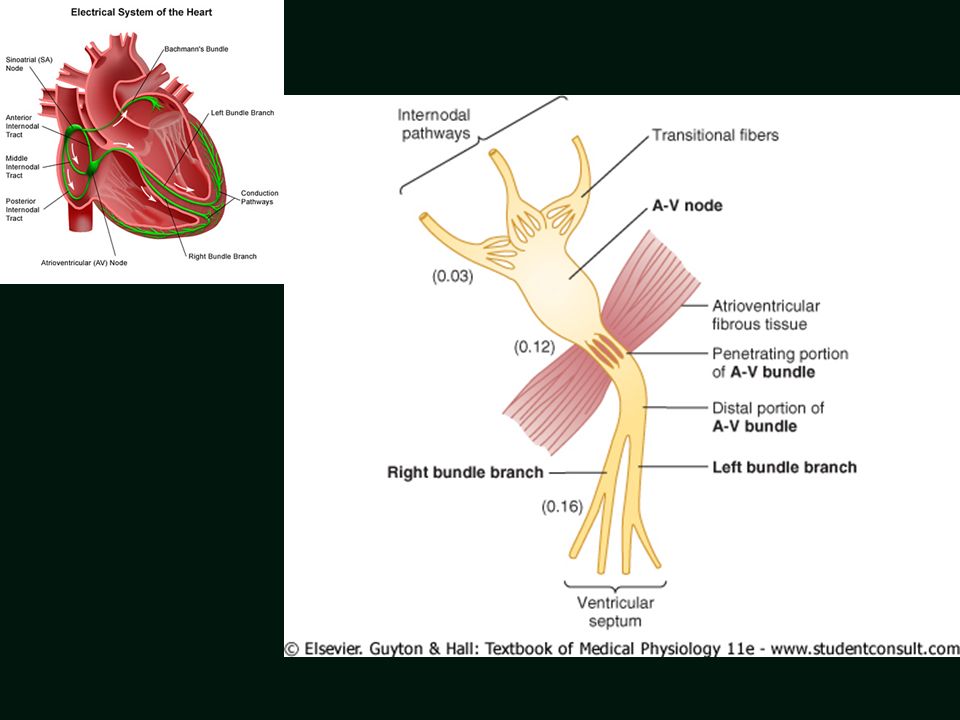
Electrical system of the heart
3 possible Pacemakers – Primary – Sinoatrial node, Secondary – Atrioventricular node Tertiary – Purkinje fibers Atrioventricular node 46

48
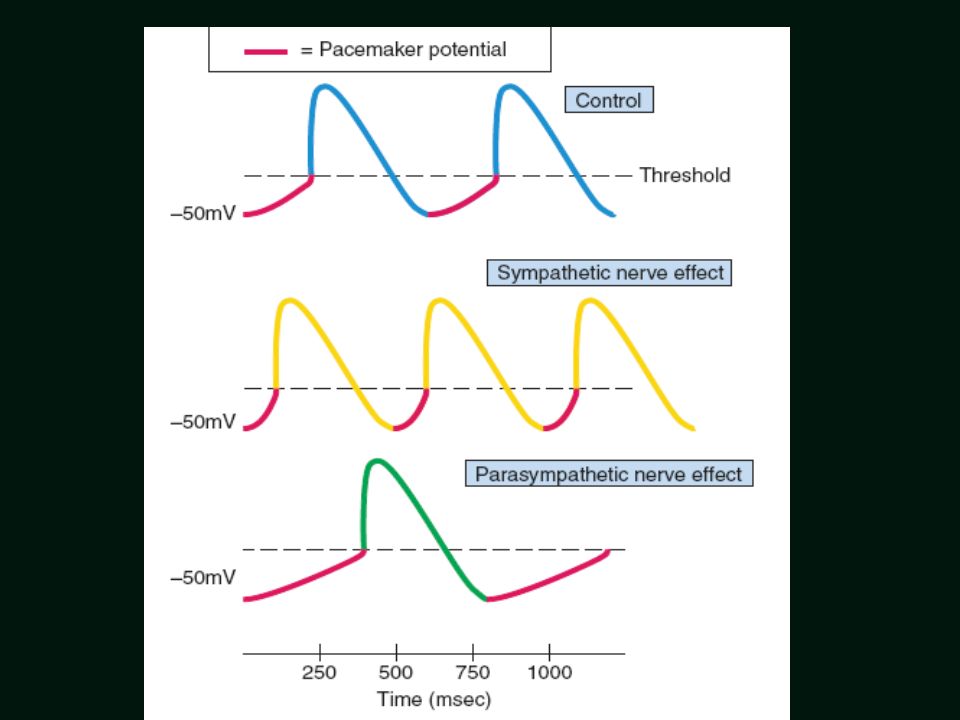
Własny rytm rozrusznikowy:
ROZRUSZNIK PIERWSZORZĘDOWY Węzeł zatokowy = 60 – 90/min ROZRUSZNIK DRUGORZĘDOWY Węzeł przedsionkowo-komorowy = 40 – 60/min ROZRUSZNIKI TRZECIORZĘDOWE Włókna Purkinjego = /min Kardiomiocyty komór serca < 30/min (5 – 30/min) 48
 48.")
49
ICa+ IK+ INa+ If Na+/K+ MAJOR MYOCYTE ION CHANNELS (activated during
depolarization) repolarizing (all myocytes) rapid depolarizing (non-nodal) depolarizing (nodal AP and myocyte contraction) Read- Table 20-1 Na+/K+ “funny channel” or HCN Pacemaker current (activated during hyperpolarization) Hyperpolarization activated Cyclic Nucleotide gated channel Which channel is absent in SA and AV node? Absent in ventricular myocytes? If 49
 repolarizing. (all myocytes) rapid. depolarizing. (non-nodal) depolarizing. (nodal AP. and myocyte. contraction) Read- Table Na+/K+ funny channel or HCN. Pacemaker current. (activated during hyperpolarization) Hyperpolarization activated Cyclic. Nucleotide gated channel. Which channel. is absent in SA and AV node Absent in ventricular myocytes If. 49.")
52
Electrical system of the heart
3 possible Pacemakers – Primary – Sinoatrial node, Secondary – Atrioventricular node Tertiary – Purkinje fibers Atrioventricular node 52

53
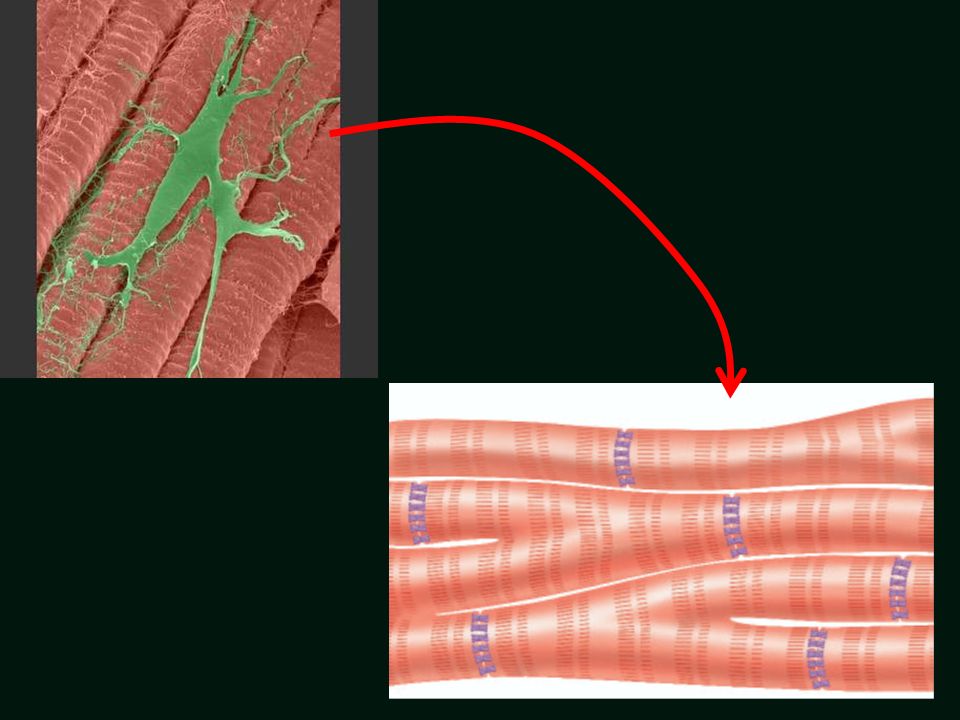
Włókna Purkiniego na tle typowych włókien mięśniowych w sercu.
Włókna Purkiniego na tle typowych włókien mięśniowych w sercu. Włókna Purkiniego są zmodyfikowanymi kardiomiocytami, które wywodzą się przede wszystkim z węzła przedsionkowo-komorowego i które rozprzestrzeniły się do obu komór. Przewodzą pobudzenie do włókien roboczych komór, zapewniając szybki i zsynchronizowany skurcz. Włókna Purkiniego przewodzą pobudzenie niezwykle szybko – do 4 m/s.

55
Cell to cell communication in heart

57
Przewodzenie pobudzenia przez złącza ścisłe w mięśniówce serca
Przyczyną przepływu fali pobudzenia jest różnica potencjału (ΔVAB) pomiędzy sąsiadującymi komórkami A i B W momencie pobudzenia komórki A, komórka B nie jest jeszcze pobudzona → różnica potencjałów (ΔVAB) 3. Kardiomiocyty są nieszczelne. Po aktywacji (powstaniu potencjału czynnościowego) komórki A i po przepłynięciu kationów (Ca2+, Na+) przez złącza ścisłe z komórki A do komórki B, kationy te z jednej strony depolaryzują komórkę, z drugiej zaś wypływają z komórki przez otwarte kanały jonowe i wymienniki jonowe (np. Na/Ca).
 pomiędzy sąsiadującymi komórkami A i B. W momencie pobudzenia komórki A, komórka B nie jest jeszcze pobudzona → różnica potencjałów (ΔVAB) 3. Kardiomiocyty są nieszczelne. Po aktywacji (powstaniu potencjału czynnościowego) komórki A i po przepłynięciu kationów (Ca2+, Na+) przez złącza ścisłe z komórki A do komórki B, kationy te z jednej strony depolaryzują komórkę, z drugiej zaś wypływają z komórki przez otwarte kanały jonowe i wymienniki jonowe (np. Na/Ca).")
58
Przewodzenie depolaryzacji podprogowej przez złącza ścisłe w mięśniówce serca
Jeżeli w komórce A dojdzie do depolaryzacji, która nie osiągnęła poziomu progowego dla otwarcia kanału sodowego ( np. wskutek niedotlenienia tej komórki lub otrzymania kationów z sąsiedniej komórki) to: zaistnieje różnica potencjałów między tą komórką, a komórka sąsiednią. dojdzie do przepływu ładunku (kationy) z komórki A do komórki B, a następnie, na podobnej zasadzie z komórki B na C, z C na D, z E na F itd.) Ponieważ komórka, która otrzymała kationy przez złącze ścisłe jest „nieszczelna”, część z otrzymanego ładunku uwolni się na zewnątrz. W efekcie komórka B będzie mniej elektroujemna (mniej zdepolaryzowana) od komórki A,. Podobnie komórka C będzie mniej elektroujemna od B, D od C itd. W ten sposób pobudzenie podprogowe szerzy się w mięśniówce serca z postępującą stratą (dekrementem) W efekcie jedna komórka częściowo zdepolaryzowana może spowodować heterogenność potencjału spoczynkowego w swojej okolicy.
 to: zaistnieje różnica potencjałów między tą komórką, a komórka sąsiednią. dojdzie do przepływu ładunku (kationy) z komórki A do komórki B, a następnie, na podobnej zasadzie z komórki B na C, z C na D, z E na F itd.) Ponieważ komórka, która otrzymała kationy przez złącze ścisłe jest „nieszczelna , część z otrzymanego ładunku uwolni się na zewnątrz. W efekcie komórka B będzie mniej elektroujemna (mniej zdepolaryzowana) od komórki A,. Podobnie komórka C będzie mniej elektroujemna od B, D od C itd. W ten sposób pobudzenie podprogowe szerzy się w mięśniówce serca z postępującą stratą (dekrementem) W efekcie jedna komórka częściowo zdepolaryzowana może spowodować heterogenność potencjału spoczynkowego w swojej okolicy.")
59
Zgodnie z prawem Ohma przepływ prądu ( I ) jest wprost proporcjonalny do różnicy potencjałów między komórkami (ΔVAB) , a odwrotnie proporcjonalny do oporu (R) ścisłego złącza. Przy wysokim nadstrzale fazy 0 (szczyt fazy 0 znacznie elektro-dodatni), pobudzenie rozprzestrzenia się szybko i bez przeszkód w komórkach mięśnia sercowego, bo strata szybko przemieszczającego się ładunku z komórki na komórkę jest niewystarczająca, aby pobudzenie zastopować (tzn. doprowadzić do depolaryzacji poniżej poziomu wyładowań; przy dużej szybkości przewodzenia niewiele kationów zdąży opuścić kolejne komórki). Tak się dzieje, jeżeli: potencjał czynnościowy jest pierwotnie zainicjowany z poziomu wysokiego (znacznie elektroujemnego) potencjału komórki. Jeżeli potencjał komórki jest mniejszy (komórka mniej elektroujemna) np.. wskutek niedoborów energetycznych lub pobudzenia przywspółczulnego prowadzącego do zmniejszenia szybkości przewodzenia pobudzenia z przedsionków do komór, to mogą zaistnieć warunki, w których nie wszystkie kardiomiocyty ulegną pobudzeniu (po pobudzenie szerząc się z dekrementem na pewnym odcinku zaniknie). W efekcie: kardiomiocyty w danym obszarze ściany serca stają się elektrycznie heterogenne, co stwarza warunki do powstania ognisk bodźcotwórczych. siła skurczu komory zmniejsza się, w czym udział ma niepełna synchronizacja skurczu poszczególnych kardiomiocytów
 np.. wskutek niedoborów energetycznych lub pobudzenia przywspółczulnego prowadzącego do zmniejszenia szybkości przewodzenia pobudzenia z przedsionków do komór, to mogą zaistnieć warunki, w których nie wszystkie kardiomiocyty ulegną pobudzeniu (po pobudzenie szerząc się z dekrementem na pewnym odcinku zaniknie). W efekcie: kardiomiocyty w danym obszarze ściany serca stają się elektrycznie heterogenne, co stwarza warunki do powstania ognisk bodźcotwórczych. siła skurczu komory zmniejsza się, w czym udział ma niepełna synchronizacja skurczu poszczególnych kardiomiocytów.")