Pobierz prezentację
1
Chromatyna a epigenetyka

2
Chromatyna wypełnia jądro komórkowe
Chromatyna - kompleks nukleoproteinowy DNA, histony, białka niehistonowe Funkcja strukturalna oraz regulacyjna W jadrze nie ma nagiego DNA tylko chromatyna, co wpływa decydujaco na przebieg zachodzacych w nim procesow:replikacja, transkrypcja itd… , zaburzenia wystepuja w wielu chorobach, m.in. nowotworach

3
23 ludzkie chromosomy w jądrach fibroblastów we wczesnej profazie

4
Maksymalny stopień kondensacji DNA osiąga w chromosomach
W jadrze interfazowym jest bardziej rozluzniona

5
Białka histonowe Histony-uniwersalne u eukariota (jaki jest wyjątek? - protaminy) Małe, zasadowe – lizyna i arginina Domeny globularne- rownomiernr rozmieszczenie ladunku,; odpowiedzialne za oddziaływania międzyhistonowe histonow rdzeniowych oraz wiazanie z DNA ;ogony sa zasadowe , w H1 N i C końcowe Bardzo konserwowane – najbardziej H4, 2 różnice miedzy krową i grochem. Ogony sa bardziej zmienne szczególnie w H1, H2A i H2B Różnice w sekw aa sa podstawa do wyroznienia wariantow sekwencyjnych , szczególnie duzo dla h1, brak dla H4. są kodowane przez odrebne geny.

6
Dean Hewish, 1973 Leigh Burgoyne, 1973
Chromatyna trawiona nukleazą, oryginał z pracy Hewish and Burgoyne 1973 Dean Hewish, 1973 Leigh Burgoyne, 1973

7
Trawienie chromatyny MNazą - drabinka nukleosomowa
Mono, di, tri nukleosomy itd. Dlugie trawiienie ujawnia chromatosom i cz. Rdzeniowa – coraz krotsze DNA

8
„Sznur korali” (‘beads on the string’)
Olins & Olins, 1973

9
NUKLEOSOM JEST PODSTAWOWĄ JEDNOSTKĄ STRUKTURALNĄ CHROMATYNY
Roger Kornberg w 1974 r. zaproponował model, w którym DNA owinięty jest wokół rdzenia histonowego tworząc nukleosom NUKLEOSOM JEST PODSTAWOWĄ JEDNOSTKĄ STRUKTURALNĄ CHROMATYNY aureat Nagrody Nobla w dziedzinie chemii w 2006 9

10
Fałd histonowy Charakt motyw strukturalny dla hist rdzeniowych odpowiada za interakcje histonow oktameru w nukleosomie Ok. 70aa, 3 helisy i krótkie łączniki

11
Złożenie fałdów (hand shake)
Silne oddziaływanie

12
Konserwowane elementy na obrzeżu oktameru
Odpowiedzialne za interakcje histony-DNA

13
Oktamer – oddziaływanie z DNA
Podstawową, powtarzalną jednostką strukturalną chromatyny jest nukleosom. W budowie nukleosomu można wyróżnić tzw. cząstkę rdzeniową – zwartą strukturę w kształcie spłaszczonego cylindra, złożoną z ośmiu cząsteczek histonów (po dwie cząsteczki histonów H2A, H2B, H3 i H4) na które nawinięty jest fragment DNA. Ogony wystaja na zewnatrz nukleosomu
 na które nawinięty jest fragment DNA. Ogony wystaja na zewnatrz nukleosomu.")
14
Mutacje SIN znoszą oddziaływania oktameru z DNA
SIN (SWI/SNF independent) powoduja zmiane struktury chromatyny
 powoduja zmiane struktury chromatyny.")
15
Składanie nukleosomu Nukleosom = H3H4 tetramer + 2 dimery H2AH2B
Udział czaperonow przy składaniu

16
Zaginanie i zwijanie DNA na oktamerze
Samo nawiniecie dna na oktamery wymusza pewna konformacje DNA Ściśle zwinięta lewoskrętna superhelisa ma ok. 80 pz na zwój i skok superhelikalny ok. 27,5 A.

17
Ekspozycja miejsc w helisie DNA na oktamerze
Jedne fragmenty sa skierowane do wewnatrz, inne na zewnatrz nukleosomu

18
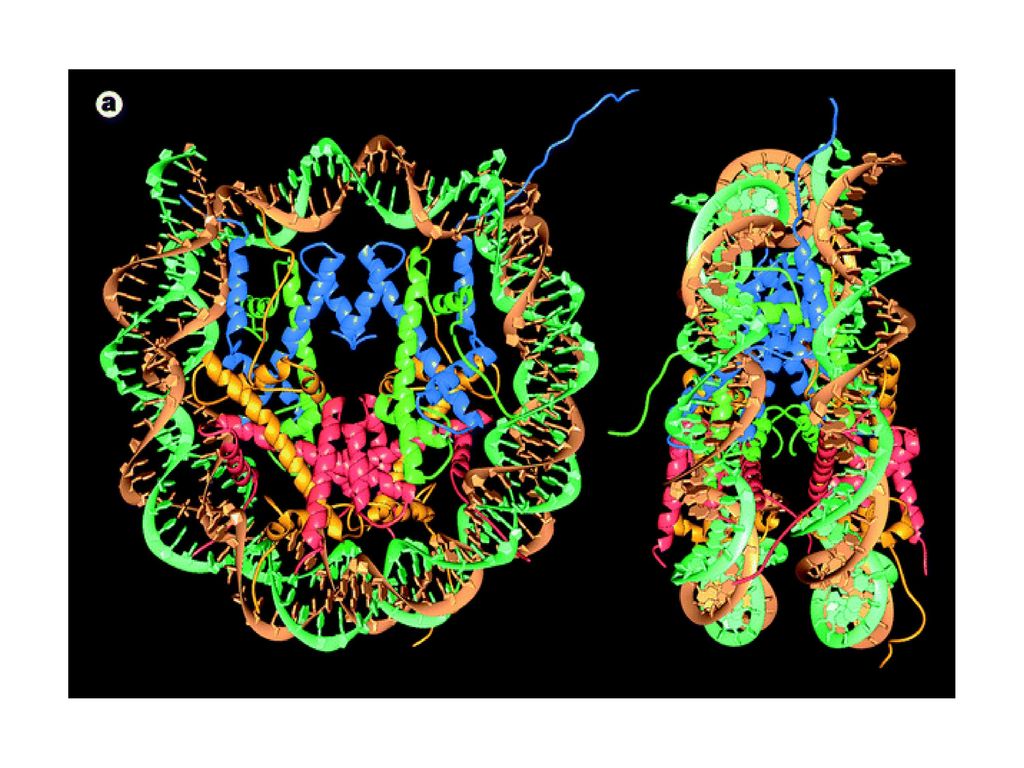
Struktura krystalogtaficzna cząstki rdzeniowej nukleosomu
Karolin Luger Timothy Richmond Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ. Nature 1997 Sep18;389(6648):251-60 18
:")
20
Schemat nukleosomu

21
Nukleosomy - terminologia
8 histonów: Po dwa każdego z: H2A H2B H3 H4 Gräff and Mansuy (2008). Behav Brain Res.
. Behav Brain Res.")
22
Organizacja chromatyny
Podwójna helisa DNA 2 nm Histony + DNA. “Koraliki na sznurku” f 11 nm Komórka roślinna Włókna nukleosomowe 30-nm (solenoid) 30 nm Włókna połączone z macierzą jądrową 300 nm Nuclear Matrix
 30 nm. Włókna połączone. z macierzą jądrową. 300 nm. Nuclear Matrix.")
23
Umiejscowienie H1 w nukleosomie
Albo wiaże się w jednym miejscu na DNA (po lewej) albo spina DNA lacznikowy w miejscu wejscia i zejscia
 albo spina DNA lacznikowy w miejscu wejscia i zejscia.")
24
Regulacyjna rola chromatyny
Struktura chromatyny nie jest jednolita, tzn. w jądrze komórkowym obok siebie występują regiony o niskim oraz wysokim stopniu kondensacji nazywane tradycyjnie eu- i heterochromatyną

25
Struktura a funkcja chromatyny
Połączone ze sobą nukleosomy tworzą w roztworach o małej sile jonowej włókno chromatynowe o średnicy 10nm, które w warunkach fizjologicznych spontanicznie fałduje się w grubsze włókno o średnicy 30nm. Nie wyjaśniono do tej pory, jak zbudowane są struktury chromatyny wyższego rzędu Rola histonu H1 i ogonów rdzeniowych Różne formy chromatyny mogą wzajemnie miedzy soba przechodzic – zmiany te sa regulowane i decydują o przebiegu procesów genetycznych (np.. Transkrypcji ) w jądrze
 w jądrze.")
26
Zmiany struktury chromatyny
modyfikacje DNA modyfikacje potranslacyjne histonów wyspecjalizowane warianty histonów ATP-zależna przebudowa (remodeling) chromatyny
 chromatyny.")
27
Metylacja DNA, Metylomy

28
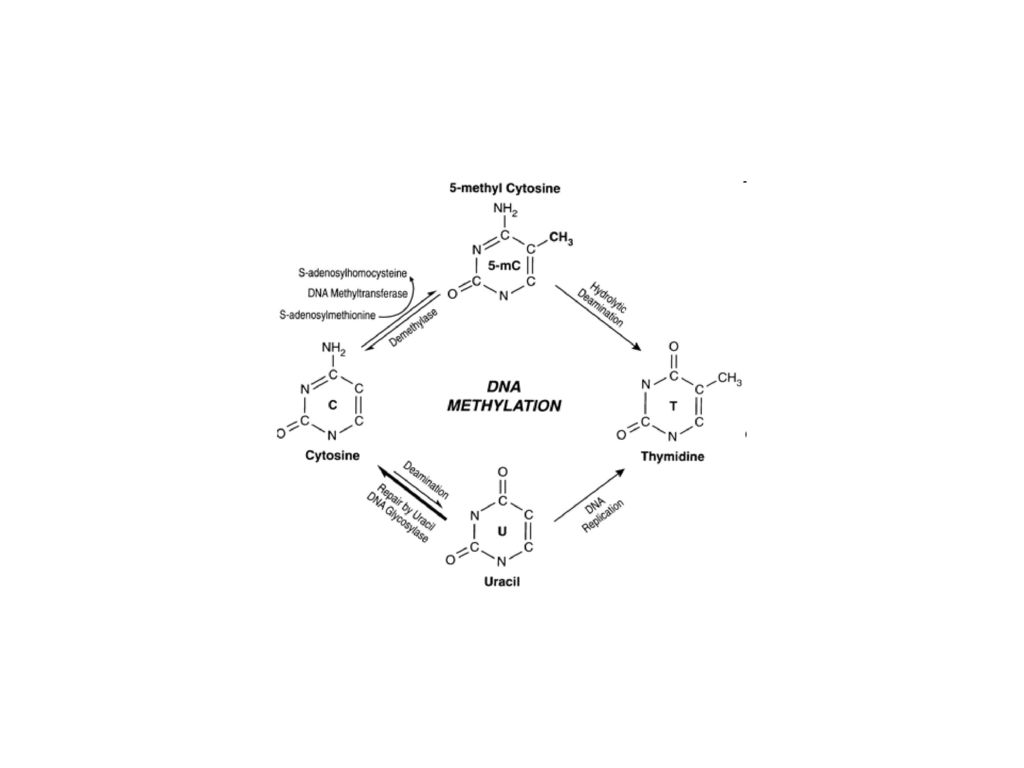
Metylacja DNA zamienia cytozynę w 5-metylo cytozynę

30
Metylacja DNA ma znaczenie biologiczne
Arabidopsis 46-dni od wysiania O N NH2 ~ CH3 cytosine 5-methylcytosine Methyltransferase WT met1/cmt3 Xiao, et al.(2006). The Plant Cell. 18:
. The Plant Cell. 18:")
31
Metylacja DNA u eukariontów
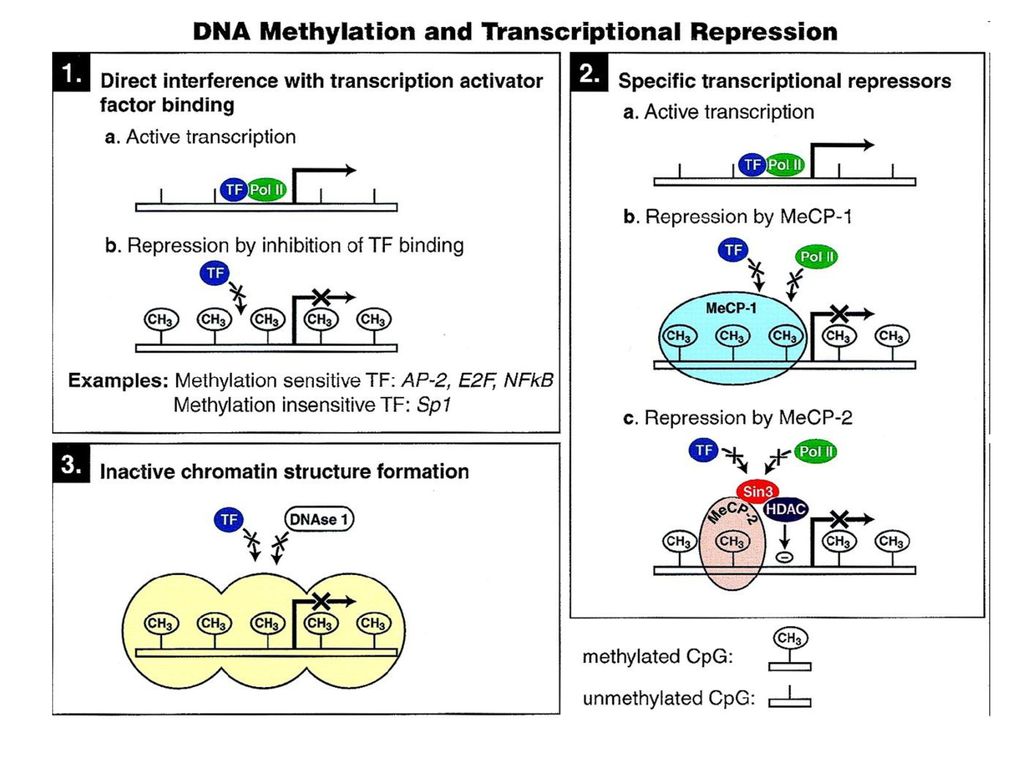
Nie jest uniwersalna, występuje u ssaków i roślin kwiatowych Jest zmienna gatunkowo, tkankowo i chromosomowo Rozpoznawana przez rodzinę białek zawierających domenę (MBD) wiążącą metylowany DNA Ściąga kompleksy białkowe indukujące zmiany w lokalnej strukturze chromosomów Wyłącza ekspresję genów Zakłócenia w normalnym wzorze metylacji DNA są niemal powszechnie wykrywane w nowotworach
 wiążącą metylowany DNA. Ściąga kompleksy białkowe indukujące zmiany w lokalnej strukturze chromosomów. Wyłącza ekspresję genów. Zakłócenia w normalnym wzorze metylacji DNA są niemal powszechnie wykrywane w nowotworach.")
33
Transpozony Fragmenty DNA, które mogą się wstawiać w nowe miejsce w chromosomie Niektóre są zdolne do autokopiowania, co prowadzi do wzrostu ich ilości w genomie Odpowiadają za wielkoskalowe zmiany w chromosomach, jak również pojedyncze wydarzenia mutacyjne

34
U roślin program epigenetyczny wycisza transpozony i chroni integralość centromerów

35
Transpozony mogą powodować wyłączenie lub niestabilność alleli
Allel dziki Ziarniak barwny Allel zmutowany Ziarniak bezbarwny Allel niestabilny Ziarniak częściowo zabarwiony Wycięcie transpozonu powoduje niestabilność allelu Gen biosyntezy barwnika Gen rozbity przez transpozon

36
Metylacja DNA jest niezbędna do wyciszania transpozonów
Brązowy = Metylowany DNA w mutancie met1 Niebieski = gęstość genów Czerwony = Gęstość elementów powtarzalnych Zielony = Metylowany DNA Utrata funkcji met1 lub ddm1 (decrease in DNA methylation1) mutanty mają niedometylowany DNA Zhang, et al. (2006). Cell 126:
 mutanty mają niedometylowany DNA. Zhang, et al. (2006). Cell 126:")
37
Metylotransferazy DNA w Arabidopsis
MET1 (METHYLTRANSFERASE1) Miejsca 5'-CG-3' Wyciszanie transpozonów, elementów powtórzonych, piętnowanie genów (imprinting) CMT3 (CHROMOMETHYLASE3) Miejsca 5'-CHG-3' (H= A, C or T) Oddziaływanie z modyfikakcjami histonów DRM 1, DRM 2 (DOMAINS REARRANGED 1 and 2) Miejsca 5'-CHH-3' Metyluje głównie elementy powtarzalne Wymaga aktywnej indukcji przez siRNA
 Miejsca 5 -CG-3 Wyciszanie transpozonów, elementów powtórzonych, piętnowanie genów (imprinting) CMT3 (CHROMOMETHYLASE3) Miejsca 5 -CHG-3 (H= A, C or T) Oddziaływanie z modyfikakcjami histonów. DRM 1, DRM 2 (DOMAINS REARRANGED 1 and 2) Miejsca 5 -CHH-3 Metyluje głównie elementy powtarzalne. Wymaga aktywnej indukcji przez siRNA.")
38
Aktywacja transpozonów w mutancie ddm1 indukuje mutacje
Po wyłączeniu DDM1, fenotyp rośliny staje się coraz bardziej nienormalny, w miarę akumulowania indukowanych przez transpozony. Dziki Po 1 pokoleniu Po 3 pokoleniach Po 5 Kakutani, et al. (1996). PNAS. 93:
. PNAS. 93:")
39
Lokalizacja ogonów histonowych w nukleosomie
H2A H2A H2B H2B

40
Enzymy modyfikujące histony i DNA
HAT – acetylotransferazy histonów (bromodomena) HDAC – deacteylazy histonów HMT – metylotransferazy histonów (chromodomena). CMT3 – DNA metylotransferaza (chromodomena)
 HDAC – deacteylazy histonów. HMT – metylotransferazy histonów. (chromodomena). CMT3 – DNA metylotransferaza (chromodomena)")
41
Przykład kowalencyjnej modyfikacji aminokwasów -acetylacja lizyny

42
Elektroforetyczny dowód acetylacji histonów

43
Efekt acetylacji ogonów histonowych
Inhibitory deacetylaz – leki przeciwnowotworowe i w chorobach OUN

44
Modyfikacje potranslacyjne białek histonowych
Kozaurides 2007

45
Modyfikacje a struktura chromatyny
H3 K9 diME H3 K9 diME euchromatyna heterochromatyna

46
Modyfikacje histonów * * * k6 Biotinylation ADP-ribosylation k11
Gräff and Mansuy (2008). Behav Brain Res.
. Behav Brain Res.")
47
Po-translacyjne modyfikacje histonów w nukleosomie wg. B
Po-translacyjne modyfikacje histonów w nukleosomie wg. B.Turner, Cell 2002

48
Mechanizm działania modyfikacji potranslacyjnych białek histonowych
Działanie bezpośrednie: zmiany w oddziaływaniach histon-DNA i histon-histon Działanie pośrednie: rekrutacja białek rozpoznających określone modyfikacje Kozaurides 2007

49
Przykład przeciwstawnych funkcji modyfikacji histonów
H3/H4 AcLys/H3metLys4(H3K4) - związane z rejonami aktywnej transkrypcji H3/H4 Lys+/H3metLys9 (H3K9) - związane z rejonami wygaszonej transkrypcji.
 - związane z rejonami aktywnej transkrypcji. H3/H4 Lys+/H3metLys9 (H3K9) - związane z rejonami wygaszonej transkrypcji.")
50
Determinanty aktywnej i wyciszonej chromatyny

51
Euchromatyna i heterochromatyna
Mniej skondensowana Głównie w ramionach chromosomów Zawiera unikatowe sekwencje Bogata w geny Silnie skondensowana Głównie w centromerach i telomerach Zawiera powtórzone sekwencje Uboga w geny

52
Znaczniki epigenetyczne związane ze stanami chromatyny
ATP-zależna przebudowa chromatyny ATP-zależna przebudowa chromatyny ATP-dependent chromatin remodeling

53
Warianty histonów – H2A Warianty h1- rozna dynamika wiazania z chromatyna

54
Warianty histonów - rozmieszczenie w chromosomach

55
SWI/SNF ISWI Mi2 Snf2 ATP-zależna przebudowa (remodeling) chromatyny
Główne typy kompleksów remodelujących chromatynę SWI/SNF ISWI Mi2 Swp73 Swi3 Snf5 Snf2 ISWI

56
Działanie kompleksów remodelujących chromatynę
Kingston & Narlikar, 1999

57
ATP-zależna przebudowa (remodeling) chromatyny
Jeden z mechanizmów remodelingu: przesunięcie oktameru histonów wzdłuż nici DNA

58
Przykład „przebudowy” (remodelingu) chromatyny
Pozycja skrajna Pozycja centralna

59
Pozycjonowanie nukleosomów

60
Nukleosomy zajmują określone miejsca w chromatynie
A. Schemat ułożenia nukleosomów w chromatynie, pojęcie długości powtórzeń nuklesomowych. B. Silnie zlokalizowane i rozmyte nukleosomy. C. Nukleosomy a miejsca wiązania czynników transkrypcyjnych. G. Arya et al. jbsd 2010

61
Nukleosomy a transkrypcja
Schemat otwartych (A) i zamkniętych (B) promotorów. Typowa mapa pozycyjna nukleosomów na otwartym promotorze pokazuje wyraźne wyróżnienie locus genowego przez obecność rejonów 5’ i 3’ wolnych od nukleosomów (NDR). (TSS) - miejsce startu transkrypcji. 5’ NDR sąsiaduje z silnie związanymi nukleosomami. G. Arya et al. jbsd 2010
 i zamkniętych (B) promotorów. Typowa mapa pozycyjna nukleosomów na otwartym promotorze pokazuje wyraźne wyróżnienie locus genowego przez obecność rejonów 5’ i 3’ wolnych od nukleosomów (NDR). (TSS) - miejsce startu transkrypcji. 5’ NDR sąsiaduje z silnie związanymi nukleosomami. G. Arya et al. jbsd")
62
Dziedziczenie epigenetyczne
Dziedziczne modyfikacje funkcji genów nie związane ze zmianami w sekwencji DNA Mechanizmy epigenetyczne są związane z modulacją chromatyny i obejmują: modyfikacje histonów, metylację DNA, przebudowę nukleosomów (remodeling), system RNAi.
, system RNAi.")
63
Epigenetyka Dziedziczenie właściwości (np. wzoru ekspresji genów) poprzez mitozę, nie wymagające zmian w sekwencji DNA. Genetyka Epigenetyka Allis, et al.(2007).Epigenetics, overview and concepts. CSHL.
.Epigenetics, overview and concepts. CSHL.")
64
Istota modyfikacji epigenetycznej

65
Dezaktywacja chromosomu X polega na wyciszeniu epigenetycznym
U samic ssaków, w każdej komórce jedna z kopii chromosmu X jest epigenetycznie wyłączona X XX U samic ssaków, w każdej komórce jedna kopia chromosomu X jest wyciszona epigenetycznie. Kolor futra u kotów jest częściowo determinowany przez gen orange, położony w chromosomie X. Samica, która jest heterozygotą w genie orange, wykazuje czarne i pomarańczowe plamy, które odpowiadają komórkom, z genem wyłączonym w jednym lub drugim chromosomie.

66
Dlaczego sklonowany kot nie jest dokładnie taki, jak oryginał?
Rainbow Klon Rainbow

67
U bliźniąt monozygotycfznych różnice epigenetyczne (metylacja DNA) nagromadzają się w życiu osobniczym Istotne różnice w metylacji DNA pokazane są jako grube czerwone i zielone rejony na ideogramach. Para 50-letnich bliźniaków wykazuje liczne zmian e we wzorze metylacji DNA (zielone=hipermetylacja; czerwone=hipometylacja), 3-letnie bliźnięta maja bardzo podobny wzór metylacji (żółte). "...Stwierdziliśmy, że choć monozygotyczne bliźnięta są epigenetycznie nierozróżnialne we wczesnych latach życia, w starszym wieku wykazują znaczące różnice w zawartości i rozkładzie 5-metylocytozyny w DNA oraz w acetylacji histonów, co wpływa na profil ekspresji ich genów. Pokazuje to jak bardzo niedoceniamy znaczenia epigenetyki w powstawaniu różnych fenotypów z tego samego genotypu." Proc Natl Acad Sci U S A Jul 26;102(30): Copyright (2005) National Academy of Sciences, U.S.A
, 3-letnie bliźnięta maja bardzo podobny wzór metylacji (żółte). ...Stwierdziliśmy, że choć monozygotyczne bliźnięta są epigenetycznie nierozróżnialne we wczesnych latach życia, w starszym wieku wykazują znaczące różnice w zawartości i rozkładzie 5-metylocytozyny w DNA oraz w acetylacji histonów, co wpływa na profil ekspresji ich genów. Pokazuje to jak bardzo niedoceniamy znaczenia epigenetyki w powstawaniu różnych fenotypów z tego samego genotypu. Proc Natl Acad Sci U S A Jul 26;102(30): Copyright (2005) National Academy of Sciences, U.S.A.")
68
Fenotyp mutantów w metylotransferazie histonów
Mutanty są pozbawione metylacji w H3K36. Dong, et al. (2008). Biochemical and Biophysical Research Communications. 373:
. Biochemical and Biophysical Research Communications. 373:")
69
Epigenetyczna kontrola czasu kwitnienia
Przedłuzony okrers chłodu Rozwój wegetatywny Rozwój generatywny Jesień Zima Wiosna Niektóre rośliny wymagają przedłużonego okresu chłodu (wernalizacji) – w trakcie zimy, zanim będą mogły zakwitnąć.
 – w trakcie zimy, zanim będą mogły zakwitnąć.")
70
FLC hamuje FT, aktywator kwitnienia
Roślina dzika FLC Gen FT Związanie FLC hamuje transkrypcję FT Roślina z mutacją flc Gen FT FT

71
Mutanty w genie FLOWERING LOCUS C (FLC) kwitna wcześnie
Jesień Zima Wiosna FLC jest inhibitorem kwitnienia; brak FLC znosi konieczność wernalizacji.

72
FLC jest wyciszany w trakcie wernalizacji
Po 40 dniach w 4°C , FLC nie jest wyrażany. 10 dni po powrocie do 22°C ekspresja FLC jest wciąż wyłączona. Jesień Zima Wiosna FLC transkrybowany FLC wyciszony Sung, et al. (2004). Nature. 427:
. Nature. 427:")
73
FLC jest regulowany przez modyfikacje epigenegtyczne
H3k4me H3K36me H3K9ac H314ac H3k9me2 H3k27me2 Zimno Jesień Zima Wiosna FLC transkrybowany FLC wyciszony

74
Kompleksy VIN3 i PRC2 epigenetycznie wyłączają FLC
PRC2= Polycomb Represive Complex 2) VIN3 PRC2 (łącznie z VIN3) Jesień Zima Wiosna FLC transkrybowany FLC wyciszony
 VIN3. PRC2 (łącznie z VIN3) Jesień. Zima. Wiosna. FLC transkrybowany. FLC wyciszony.")
75
Program epigenetyczny wspomaga przejścia rozwojowe u roślin
Przejście z fazy embrionalnej do wegetatywnej Przejście z fazy wegetatywnej w generatywną Rozwój embrionalny Rozwój wegetatywny Rozwój generatywny

76
Przykład ewolucyjnego efektu epimutacji (zmiany we wzorze metylacji DNA)
Lcyc kontroluje symetrię grzbietowo-brzuszną kwiatu; u mutanta nieaktywny z powodu silnej, dziedziczonej metylacji From Cubas et al 1999, Nature 401:





 ogół rzeczywistych jednostek, o których chcemy uzyskać informacje.>")



1806 r. - J. Berzelius wprowadził nazwę „związki organiczne” dla wszystkich substancji występujących w organizmach roślinnych.>")






 (1623-1662) Blaise Pascal Ciśnienie wywierane na ciecz rozchodzi się jednakowo we wszystkich.>")


 treści prezentowanych.>")
 Cechowanie (PN-EN )>")
