Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
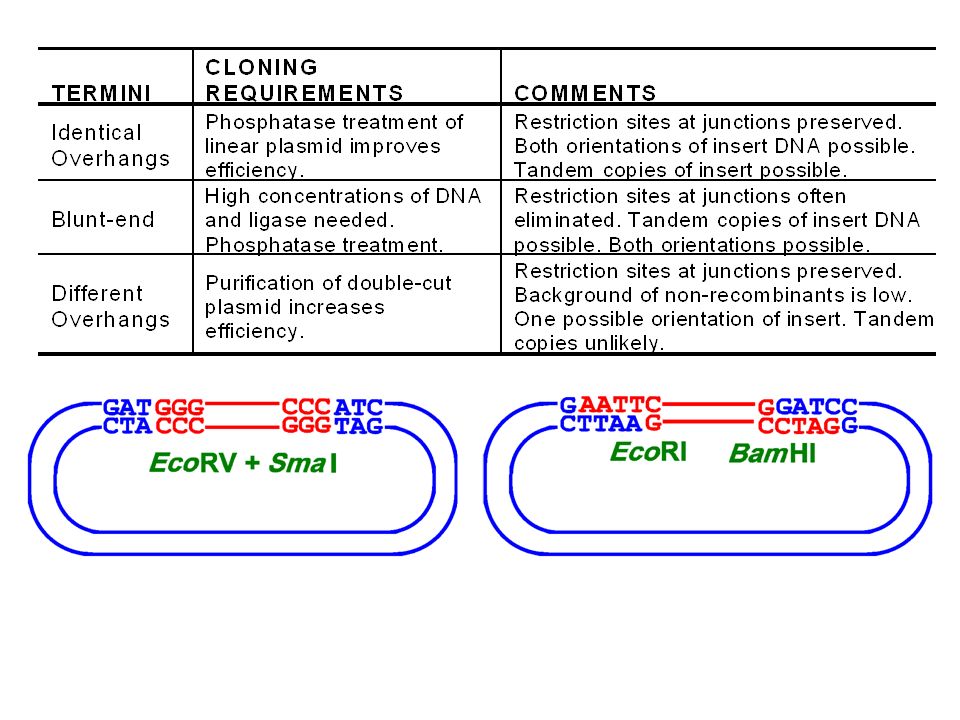
Restriction/Methylation Enzyme

2
Eco RI Restriction Enzyme
Single stranded “nick” First restriction enzyme from Escherichia coli, so Eco R1

3
Restriction Enzyme Recognition Sites
Restriction sites are general palindromic: Bam H1 site: 5’-GGATCC-3’ 3’-CCTAGG-5’

4
Restriction Enzyme Recognition Sites
All these sticky ends are compatible BglII 5’ A-G-A-T-C-T T-C-T-A-G-A 5’ Sau3A 5’ G-A-T-C C-T-A-G 5’ BamHI 5’ G-G-A-T-C-C C-C-T-A-G-G 5’ Isoschizomers: In certain cases, two or more different enzymes may recognize identical sites. (e.g. MboI also cleaves at GATC, and so is an isochizomer of Sau3A.)
")
5
Frequency of cutting of recognition enzymes
Sau 3A (GATC) cuts (¼)(¼)(¼)(¼) = once every 256 base pairs (assuming G/C = A/T, which is often does not) BamH1 (GGATCC) cuts (¼)(¼)(¼)(¼)(¼)(¼) = once every ~4Kb HindII (GTPyPuAC) cuts (¼)(¼)(½)(½)(¼)(¼) = once every ~1Kb
 cuts (¼)(¼)(¼)(¼) = once every 256 base pairs (assuming G/C = A/T, which is often does not) BamH1 (GGATCC) cuts (¼)(¼)(¼)(¼)(¼)(¼) = once every ~4Kb. HindII (GTPyPuAC) cuts (¼)(¼)(½)(½)(¼)(¼) = once every ~1Kb.")
6
“Sticky” ends + + “Blunt” ends + 5’ overhang (EcoRI)
5’-GAATTC-3’ 5’-G-OH PO4-AATTC-3’ 3’-CTTAAG-5’ ’-CTTAA-PO HO-G-5’ + 3’ overhang (PstI) 5’-CTGCAG-3’ 5’-CTGCA-OH PO4-G-3’ 3’-GACGTC-5’ ’-G-PO HO-ACGTC-5’ + “Blunt” ends 5’ overhang (SmaI) 5’-CCCGGG-3’ 5’-CCC-OH PO4-GGG-3’ 3’-GGGCCC-5’ ’-GGG-PO HO-CCC-5’ +
 5’-CTGCAG-3’ 5’-CTGCA-OH PO4-G-3’ 3’-GACGTC-5’ 3’-G-PO4 HO-ACGTC-5’ + Blunt ends. 5’ overhang (SmaI) 5’-CCCGGG-3’ 5’-CCC-OH PO4-GGG-3’ 3’-GGGCCC-5’ 3’-GGG-PO4 HO-CCC-5’ +")
7
Ligation of compatible sticky ends
Human DNA cleaved with EcoRI Corn DNA cleaved with EcoRI + 5’-C-G-G-T-A-C-T-A-G-OH 3’-G-C-C-A-T-G-A-T-C-T-T-A-A-PO4 PO4-A-A-T-T-C-A-G-C-T-A-C-G-3’ HO-G-T-C-G-A-T-G-C-5’ 5’-A-C-G-G-T-A-C-T-A-G A-A-T-T-C-A-G-C-T-A-C-G-3’ 3’-T-G-C-C-A-T-G-A-T-C-T-T-A-A G-T-C-G-A-T-G-C-5’ Complementary base pairing + DNA Ligase, + rATP recombinant DNA molecule 5’-A-C-G-G-T-A-C-T-A-G-A-A-T-T-C-A-G-C-T-A-C-G-3’ 3’-T-G-C-C-A-T-G-A-T-C-T-T-A-A-G-T-C-G-A-T-G-C-5’

8
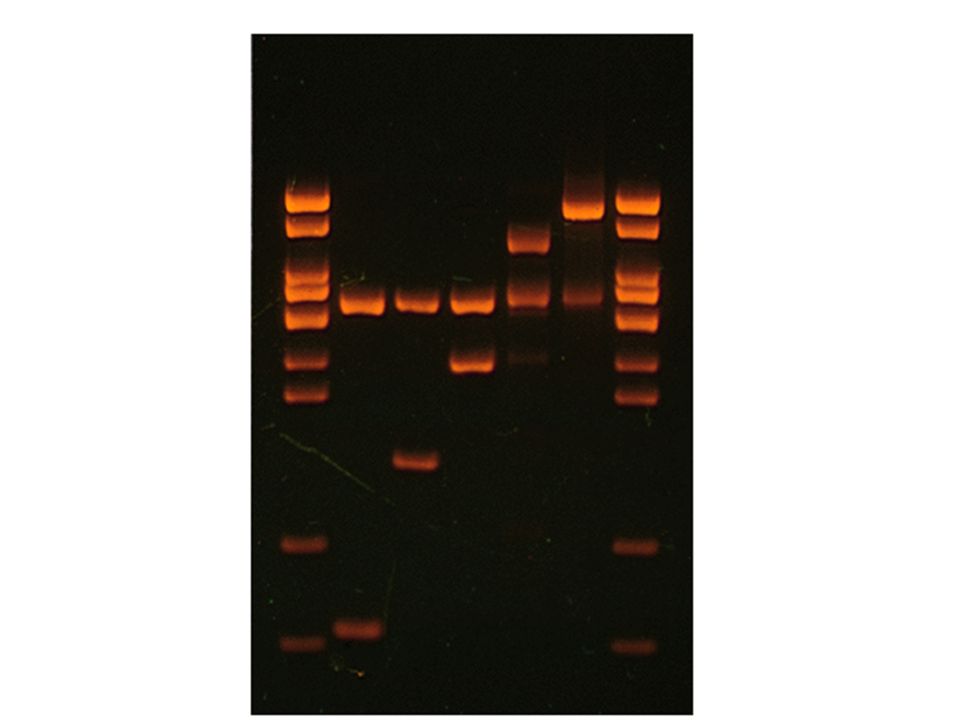
_ + Agarose Gel Electrophoresis
DNA is negatively charged from the phosphate backbone Agarose mesh + Visualize DNA with ethidium bromide – fluoresces orange ONLY when bound to DNA

10
Enzymy restrykcyjne izolowane z różnych gatunków bakterii podzielono na trzy klasy (I, II i III) w zależności od: Liczby i organizacji wchodzących w ich skład podjednostek Wymagań dotyczących kofaktorów Mechanizmu enzymatycznego Specyficzności rozpoznawanej sekwencji Regulacji ekspresji genów kodujących enzymy Podział ten jest stale modyfikowany, uaktualniany w związku z odkrywaniem nowych systemów RM

11
Typ I RM Jest najbardziej skomplikowanym systemem, złożonym z trzech podjednostek strukturalno-funkcjonalnych: Podjednostka S – rozpoznaje sekwencję DNA Podjednostka M – modyfikuje DNA Podjednostka R – aktywność restrykcyjna Podjednostki S i M tworzą m6A-metylazę DNA o stechiometrii M2S1, która rozpoznaje i modyfikuje DNA w obrębie określonej sekwencji Kompleks 3 podjednostek R2M2S1 jest enzymem restrykcyjnym (wymaga ATP) gdy napotka niezmodyfikowany DNA Cięcie następuje w różnych niezdefiniowanych odległościach od miejsca rozpoznania, zwykle kilkaset do kilku tysięcy par zasad
 gdy napotka niezmodyfikowany DNA. Cięcie następuje w różnych niezdefiniowanych odległościach od miejsca rozpoznania, zwykle kilkaset do kilku tysięcy par zasad.")
12
Typ II RM W zdefiniowanych warunkach posiadają wysoką specyficzność rozpoznawanej sekwencji Dają powtarzalne produkty trawienia endonukleolitycznego Nie wymagają ATP i S-adenozylo-L-metioniny, a jedynie jonów Mg Aktywności metylazy i endonukleazy rozdziolone są między dwa odrębne białka kodowane przez różne geny Rozpoznają krótkie najczęściej palindromiczne sekwencje 4-8 pz i trawią DNA w obrębie sekwencji rozpoznania lub w pewnej ściśle określonej odległości od niej Wyróżnia się podtypy lub klasy w obrębie rodziny II

13
Klasy typu II RM II S (monomery w roztworze; rozpoznają asymetryczną sekwencję; cięcie w zdefiniowanej odległości od sekwencji rozpoznawanej, 1-20 pz; np. FokI – GGATGN9/13 II E (rozpoznawane dwie sekwencje: w efektorze allosterycznym i właściwej sekwencji ciętej, np. NaeI – GCG/CGC) II F (homotetramer, rozpoznaje dwie sekwencje, trawienie jednoczesne obu miejsc, np. NgoMIV – G/CCGGC II T (heterodimer, rozpoznawana sekwencja asymetryczna, sekwencja palindromiczna, np. Bpu10I – CC/TNAGG
 II F (homotetramer, rozpoznaje dwie sekwencje, trawienie jednoczesne obu miejsc, np. NgoMIV – G/CCGGC. II T (heterodimer, rozpoznawana sekwencja asymetryczna, sekwencja palindromiczna, np. Bpu10I – CC/TNAGG.")
14
Klasy typu II RM c.d. II G (aktywność R i M w jednym łańcuchu polipeptydowym, cięcie poza sekwencją rozpoznania, stymulacja przez SAM, np. Eco57I - CTGAAGN14/16) II B (trawienie po obu stronach rozpoznawanej sekwencji, aktywność R i M w jednym Łańcuchu polipeptydowym, np. BcgI – NN/N10CGAN6TGCN10/NN) II M (rozpoznawana sekwencja zmetylowana, np. DpnI – GmA/TC)
 II B (trawienie po obu stronach rozpoznawanej sekwencji, aktywność R i M w jednym Łańcuchu polipeptydowym, np. BcgI – NN/N10CGAN6TGCN10/NN) II M (rozpoznawana sekwencja zmetylowana, np. DpnI – GmA/TC)")
15
Typ III RM Zbudowany z 2 podjednostek: M – modyfikującej i R – restrykcyjnej, występujących w stechiometrii R2M2 Enzymy rozpoznają 5-6 pz, nie wykazujące symetrii wewnętrznej i trawią w odległości około 25 pz od miejsca rozpoznania Wymagają do aktywności ATP i S-adenozylo-L-metioniny

16
Enzymy użyteczne w analizie restrykcyjnej DNA. Podział na grupy.
enzymy należące do grupy "regularnych 6t-ek", np.: EcoRI, BamHI, BglII, PstI, HindIII enzymy należące do grupy rozpoznających kilka specyficzności, np.: HincII - GT[PyPu]AC, AccI, AvaII, AflIII enzymy należące do grupy rozpoznających nieciągłe sekwencje palindromowe, np.: BglI (GCCNNNN'NGGC), BstXI (CGANNNNN'NTGG) enzymy rozpoznające specyficzną sekwencję, lecz przecinające DNA poza nią ("shiftery", klasa lI S), np.: FokI GGATGNNNNNNNNN(9)’ CCTACNNNNNNNNNNNNN(13)’ MboII GAAGANNNNNNNN(8)’ CTTCTNNNNNNN(7)’
, BstXI (CGANNNNN NTGG) enzymy rozpoznające specyficzną sekwencję, lecz przecinające DNA poza nią ( shiftery , klasa lI S), np.: FokI GGATGNNNNNNNNN(9)’ CCTACNNNNNNNNNNNNN(13)’ MboII GAAGANNNNNNNN(8)’ CTTCTNNNNNNN(7)’")
17
Grupy enzymów pozostawiających komplementarne końce
Istnieją grupy enzymów rozpoznające odmienne sekwencje lecz pozostawiające po trawieniu komplementarne końce DNA. Są one dostępne komercyjnie i dają duże możliwości w opracowaniu strategii wieloetapowego klonowania.

18
Grupy enzymów pozostawiających komplementarne końce
Najpopularniejsze pary enzymów pozostawiających komplementarne końce to: BamHI G’GATC’C BglII A’GATC’T BstYI (XhoII) G/A’GATC’C/T BclI T’GATC’A Sau3AI ‘GATC’
 G/A’GATC’C/T. BclI T’GATC’A. Sau3AI ‘GATC’")
19
Grupy enzymów pozostawiających komplementarne końce
XbaI T’CTAG’A NheI G’CTAG’C SpeI A’CTAG’T AvrII C’CTAG’G SalI G’TCGA’C PaeR7I C’TCGA’G XhoI C’TCGA’G AvaI C’C/TCGG/A’G

20
Izoschizomery to enzymy pochodzące z różnych organizmów bakteryjnych, ale rozpoznające taką samą sekwencję i przecinające ją identycznie. Np.: SphI GCATG’C BbuI GCATG’C

21
Neoschizomery rozpoznają taką samą sekwencję DNA lecz przecinają ją w inny sposób. Uniemożliwia to łączenie (ligację) końców, więc podczas planowania ligacji nie można tego przeoczyć! Np.: Acc65I G’GTACC KpnI GGTAC’C SmaI CCC’GGG XmaI C’CCGGG BbeI GGCGC’C EheI GGC’GCC KasI G’GCGCC NarI GG’CGCC
 końców, więc podczas planowania ligacji nie można tego przeoczyć! Np.: Acc65I G’GTACC. KpnI GGTAC’C. SmaI CCC’GGG. XmaI C’CCGGG. BbeI GGCGC’C. EheI GGC’GCC. KasI G’GCGCC. NarI GG’CGCC.")
22
Problem dam/dcm metylacji DNA w komórkach E. coli
Jeżeli sekwencje GATC lub CC A/T GG są częścią sekwencji rozpoznawanej, bądź enzym rozpoznaje i przecina taką sekwencję bezpośrednio, to fakt ten ma swoje konsekwencje, jeśli DNA jest otrzymywane w komórkach E. coli dam+ dcm+. W związku z metylacją adeniny w sekwencji GATC (dam) i wewnętrznej cytozyny w sekwencji CC A/T GG (dcm) w dzikich szczepach E. coli, należy brać pod uwagę wrażliwość danego enzymu restrykcyjnego na tego typu metylację substratu. Jeżeli zatem sekwencje GATC lub CC A/T GG nakładają się na sekwencje rozpoznawane, należy wiedzieć, że nie każde istniejące na danym DNA miejsce restrykcyjne dla odpowiedniego enzymu będzie cięte.
 i wewnętrznej cytozyny w sekwencji CC A/T GG (dcm) w dzikich szczepach E. coli, należy brać pod uwagę wrażliwość danego enzymu restrykcyjnego na tego typu metylację substratu. Jeżeli zatem sekwencje GATC lub CC A/T GG nakładają się na sekwencje rozpoznawane, należy wiedzieć, że nie każde istniejące na danym DNA miejsce restrykcyjne dla odpowiedniego enzymu będzie cięte.")
23
Problem dam/dcm metylacji DNA w komórkach E. coli
Lista niektórych enzymów, których aktywność jest zależna od zmetylowanego DNA w sekwencji dam (podkreślono). Enzymy te nie trawią takich sekwencji: BclI TGAMTCA ClaI gAMTCGAT DpnII, MboI GAMTC HphI GGTGAMtc MboII GAAGAMtc NruI TCGCGAMtc XbaI TCTAGAMtc
. Enzymy te nie trawią takich sekwencji: BclI TGAMTCA. ClaI gAMTCGAT. DpnII, MboI GAMTC. HphI GGTGAMtc. MboII GAAGAMtc. NruI TCGCGAMtc. XbaI TCTAGAMtc.")
24
Problem dam/dcm metylacji DNA w komórkach E. coli
Lista enzymów dam niezależnych (niewrażliwych): BamHI GGAMTCC BglII AGAMTCT BstYI (A/G)GAMTC(C/T) PvuI CGAMTCG Sau3AI GAMTC
: BamHI GGAMTCC. BglII AGAMTCT. BstYI (A/G)GAMTC(C/T) PvuI CGAMTCG. Sau3AI GAMTC.")
25
Problem dam/dcm metylacji DNA w komórkach E. coli
Lista enzymów dcm zależnych - wrażliwych (sekwencje dcm podkreślono): AvaII GG(A/T)CCM(a/t)gg BalI TGGCCMAgg EcoRII CCM(A/T)GG Sau96I GGNCCM(a/t)gg StuI AGGCCMTgg
: AvaII GG(A/T)CCM(a/t)gg. BalI TGGCCMAgg. EcoRII CCM(A/T)GG. Sau96I GGNCCM(a/t)gg. StuI AGGCCMTgg.")
26
Problem dam/dcm metylacji DNA w komórkach E. coli
Lista enzymów niewrażliwvch na dcm metylację: BglI GCCM(A/T)GGNNGGC BstNI CCM(A/T)GG HaeIII GGCCM(a/t)gg KpnI GGTACCM(a/t)gg NarI GGCGCCM(a/t)gg
GGNNGGC. BstNI CCM(A/T)GG. HaeIII GGCCM(a/t)gg. KpnI GGTACCM(a/t)gg. NarI GGCGCCM(a/t)gg.")
27
Warunki trawienia a niespecyficzna aktywność (ang. star activity)
Jeżeli przeprowadza się trawienie w warunkach znacznie odbiegających od optymalnych dla danego enzymu, to często zdarza się, że obserwujemy niespecyficzne cięcia. Dzieje się tak, ponieważ w tych warunkach enzym rozpoznaje sekwencje różniące się od sekwencji specyficznej (kanonicznej), np. o jedną zasadę. W przypadku EcoRI, dla którego sekwencją kanoniczną jest GAATTC takie zmienione przecinane sekwencje to np. CAATTC, GAATTG, GTATTC itp.
, np. o jedną zasadę. W przypadku EcoRI, dla którego sekwencją kanoniczną jest GAATTC takie zmienione przecinane sekwencje to np. CAATTC, GAATTG, GTATTC itp.")
28
Warunki trawienia a niespecyficzna aktywność (ang. star activity)
Do czynników mogących wywołać rozluźnioną specyficzność enzymu zalicza się: 1. stężenie glicerolu powyżej 5% 2. obecność DMSO, etanolu, glikolu etylenowego 3. zbyt niską siłę jonową mieszaniny reakcyjnej 4. zbyt wysokie pH 5. obecność innych niż Mg2+ jonów dwuwartościowych metali (np.: Mn, Cu, Zn, Fe, Co, Ca) 6. zbyt wysokie stężenie enzymu w próbce.
 6. zbyt wysokie stężenie enzymu w próbce.")
29
Tworzenie nowych miejsc restrykcyjnych przez łączenie końców DNA
Nowe miejsca restrykcyjne w zrekombinowanym DNA mogą powstawać w miejscu ligacji naturalnych końców DNA bądź po wypełnieniu ich fragmentem Klenowa polimerazy DNA I: powstanie nowego miejsca przypadkowo: SspI/ClaI (Klenow) - AAT/CGAT odtworzone ClaI (ATCGAT) ClaI (Klenow)/SspI - ATCG/ATT odtworzone ClaI EcoRV/ClaI (Klenow) - GAT/CGAT odtworzone ClaI dam-zależne (gATCGAT) EcoRI (Klenow)/PvuII - GAATT/CTG odtworzone EcoRI (GAATTC)
 - AAT/CGAT. odtworzone ClaI (ATCGAT) ClaI (Klenow)/SspI - ATCG/ATT. odtworzone ClaI. EcoRV/ClaI (Klenow) - GAT/CGAT. odtworzone ClaI dam-zależne (gATCGAT) EcoRI (Klenow)/PvuII - GAATT/CTG. odtworzone EcoRI (GAATTC)")
30
Tworzenie nowych miejsc restrykcyjnych przez łączenie końców DNA
powstanie nowego miejsca w sposób zamierzony: (np. w sytuacji konieczności zlikwidowania starego miejsca restrykcyjnego i wykreowanie nowego [uzupełnianie fragmentem Klenowa polimerazy DNA I]: EcoRI/EcoRI - GAATT/AATTC XmnI (GAANNNNTTC) HindIII/HindIII - AAGCT/AGCTT NheI (GCTAGC) TaqI/TaqI - TCG/CGA NruI (TCGCGA)
 HindIII/HindIII - AAGCT/AGCTT. NheI (GCTAGC) TaqI/TaqI - TCG/CGA. NruI (TCGCGA)")
31
Lista enzymów modyfikujących niezbędnych do klonowania
zestaw enzymów restrykcyjnych fragment Klenowa polimerazy DNA I - tworzenie tępych końców przez wypełnianie cofniętych końców 3' polimeraza DNA T4 - tworzenie tępych końców przez usuwanie jednoniciowych końców 3' lub wypełnianie cofniętych końców 3' kinaza polinukleotydowa T4 - fosforylacja końców 5' ligaza DNA T4 - łączenie końców DNA nukleaza mung bean - tworzenie tępych końców przez usuwanie jednoniciowych lepkich końców RNazaA - degradacja RNA alkaliczna fosfataza - usuwanie grup fosforanowych z końców 5'

32
What is a Cloning Vehicle or Vector?
A vector is a DNA molecule in which a foreign piece of DNA can be inserted without loss of the vector’s ability to self replicate. A vector may be a plasmid, cosmid, artificial yeast chromosome, or virus.

33
Essential Features of Plasmids
Most vectors currently in use carry a replicon from pMB1 or Col E1 Both pMB1 and ColE1 are large plasmids obtained from clinical isolates of E. coli These maintain copies in a cell pMB1 and Col E1 replicons or origin of replication ori were found in clinical isolates of E. coli. FOR MORE INFO... Hershfield, V., H. W. Boyer, C. Yanofsky, M. A. Lovett, and D. R. Helinski Plasmid ColE1 as a molecular vehicle for cloning and amplification of DNA. Proc. Natl. Acad. Sci. 71:3455

34
Essential Features of Plasmids
Plasmids such as pMB1 and ColE1 (multicopy plasmids) replicate in a “relaxed” fashion Replication functions are not encoded on the plasmid Uses functions from the host genes DNA polymerases I and III, DNA-dependent RNA polymerase, etc. The result of relaxed, versus controlled replication, is that the plasmids are maintained in high copy number.
 replicate in a relaxed fashion. Replication functions are not encoded on the plasmid. Uses functions from the host genes. DNA polymerases I and III, DNA-dependent RNA polymerase, etc. The result of relaxed, versus controlled replication, is that the plasmids are maintained in high copy number.")
35
Importance of “relaxed” replication
Plasmids can function in the absence of ongoing protein synthesis = absence of host replication If you inhibit protein synthesis with antibiotics (chloramphenicol, spectinomycin)………… These antibiotics bind to ribosomes and hence, shut down protein synthesis. Since DNA replication requires polymerase (long half life), DNA synthesis continues while cell growth does not. FOR MORE INFO... Staudenbauer, W. L Structure and replication of the colicin E1 plasmid. Curr. Top. Microbiol. Immunol. 83:93.
………… These antibiotics bind to ribosomes and hence, shut down protein synthesis. Since DNA replication requires polymerase (long half life), DNA synthesis continues while cell growth does not. FOR MORE INFO... Staudenbauer, W. L Structure and replication of the colicin E1 plasmid. Curr. Top. Microbiol. Immunol. 83:93.")
36
Importance of “relaxed” replication
Host chromosome replication is prevented Plasmids carrying the pMB1 or ColE1 replicon will continue to replicate Two to three thousand copies per cell Why is this important? FOR MORE INFO... Clewell, D. B Nature of Col E1 plasmid replication in Escherichia coli in the presence of chloramphenicol. J. Bacteriol. 110:667.

37
Rop protein (63 amino acids)
pMB1 and ColE1 replicons Direction of DNA replication RNAse H Processing RNA II ori RNA II is the RNA primer that is required for DNA synthesis RNA I is transcribed from the opposite strand and along with the Rop protein to prevent RNA II from forming a cloverleaf structure that necessary for forming stable DNA-RNA hybrids between RNA II and the plasmid DNA (negative regulation of DNA synthesis) RNAse H processes the RNA II to make it precise for DNA replication RNA I rop gene Rop protein (63 amino acids)
 RNAse H processes the RNA II to make it precise for DNA replication. RNA I. rop gene. Rop protein (63 amino acids)")
38
pMB1 and ColE1 Replicons So what will happen if we alter RNA I or rop?
Decrease negative regulation of RNA II More RNA II available More plasmid replication Example pUC plasmids have a single mutation (G->A) one nucleotide upstream of the initiation of RNA I. pUC plasmids have copies per cell
 one nucleotide upstream of the initiation of RNA I. pUC plasmids have copies per cell.")
39
Replicons Also Control Plasmid Compatibility
Plasmid compatibility – the ability of two different plasmids to co-exist in the same host Plasmids that utilize the same replication system cannot co-exist in the same bacterial cell Plasmids carrying the same replicon belong to the same incompatibility group

40
Over 30 Incompatibility Groups are Known
Plasmid Replicon Copy Number pBR 322 and its derivatives pMB1 15-20 pUC vectors pACYC and its derivatives p15A 10-12 pSC101 and its derivatives pSC101 ~5 ColE1 Remember pSC101 is a derivative of R6-5. Positively controlled by it own protein. Hence, cannot amplify with chloramphenicol

41
Plasmid Safety Some naturally-occurring plasmids can be transferred to new hosts by conjugation Conjugation requires three elements A trans-acting mobilization gene (mob) A cis-acting element (bom) A specific site that is nicked by mob (nic) Mob are codes for proteins that enable conjugation. Bom stands for basis of mobility. These plus the ori are tra genes
 A cis-acting element (bom) A specific site that is nicked by mob (nic) Mob are codes for proteins that enable conjugation. Bom stands for basis of mobility. These plus the ori are tra genes.")
42
Plasmid Safety Some of the older plasmids such as pBR322 are missing mob Some of the newer vectors such as pUC are missing the nic/bom and cannot be mobilized

43
In the 1970’s and 1980’s The first cloning vectors such as pSC101 had limited functionality The next trend was to develop smaller plasmids Advantages Increased efficiency of transformation Easier to restriction map Higher copy numbers

44
Plasmid vectors Circular DNA molecules that replicate independently of E. coli chromosome. Are present in various copy per cell - Some are very high copy (can be > 100 per cell); Others are low copy (1-25 per cell). Three key features of plasmid vectors: 1) Origin of replication (e. g. ColE1, very high copy copies per cell). 2) Antibiotic resistance (or other selectable marker). 3) Multiple cloning site (often embedded in a LacZ reporter for ease of selecting inserts)
; Others are low copy (1-25 per cell). Three key features of plasmid vectors: 1) Origin of replication (e. g. ColE1, very high copy 500 copies per cell). 2) Antibiotic resistance (or other selectable marker). 3) Multiple cloning site (often embedded in a LacZ reporter for ease of selecting inserts)")
45
Desirable features of plasmids used in cloning DNA
1. Small size (<15 kb) so that it is easier to do transformation 2. Choice of restriction sites into which to insert a fragment 3. One to several selectable markers for identifying which bacteria have “picked up” the desired plasmid with fragment
 so that it is easier to do transformation. 2. Choice of restriction sites into which to insert a fragment. 3. One to several selectable markers for identifying which bacteria have picked up the desired plasmid with fragment.")
46
Plasmids - Vectors for Cloning DNA Fragments
Plasmid - small (1 kb to 500 kb) extra-genomic, circular piece of DNA maintained by bacteria and replicated (passed on) during cell division R plasmids - carry resistance to antibiotics F plasmids - carry genes that allow transfer to other bacteria high copy number plasmids - 10 to 100 copies in a single bacterium low copy number plasmids - 1 to 4 copies in a single bacterium incompatible plasmids - cannot exist together in the same bacterium narrow-host-range plasmids - small number of species can replicate broad-host-range plasmids - many species can replicate
 extra-genomic, circular piece of DNA maintained by bacteria and replicated (passed on) during cell division. R plasmids - carry resistance to antibiotics. F plasmids - carry genes that allow transfer to other bacteria. high copy number plasmids - 10 to 100 copies in a single bacterium. low copy number plasmids - 1 to 4 copies in a single bacterium. incompatible plasmids - cannot exist together in the same bacterium. narrow-host-range plasmids - small number of species can replicate. broad-host-range plasmids - many species can replicate.")
47
The First cloning experiment involving a recombinant DNA
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. The First cloning experiment involving a recombinant DNA assembled in vitro Boyer and Cohen

48
Example 1: pBR322 as a vector used to clone fragments
4,361 bp Ampr gene (w/ PstI site) Tetr gene (w/ BamHI, , HindIII, and Sal I sites) EcoRI site not within any coding region funtions only in E. coli high copy number easily transferred to other E. coli
 Tetr gene (w/ BamHI, , HindIII, and Sal I sites) EcoRI site not within any coding region. funtions only in E. coli. high copy number. easily transferred to other E. coli.")
49
pBR322 restriction map

50
Cloning foreign DNA using the PstI site of pBR322
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Cloning foreign DNA using the PstI site of pBR322

51
Copyright © The McGraw-Hill Companies, Inc
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

52
Joining of vector to insert a) Mechanism of DNA ligase
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Joining of vector to insert a) Mechanism of DNA ligase Ligase reacts with AMP donor either ATP or NAD Activated ligase Activated ligase donates the AMP to the free 5'–phosphate creating a high energy diphosphate group With energy provided by cleavage of the bond between phosphate groups, a new phosphodiester bond is formed
 Mechanism of DNA ligase. Ligase reacts with AMP donor either ATP or NAD. Activated ligase. Activated ligase donates the AMP to the free 5 –phosphate. creating a high energy diphosphate group. With energy provided by cleavage of the bond between. phosphate groups, a new phosphodiester bond is formed.")
53
Joining of vector to insert
b) Alkaline phosphatase action
 Alkaline phosphatase action.")
54
incubate ligation mixture with ‘competent cells’
Generic rDNA Protocol prepare foreign DNA prepare vector ligate foreign DNA and vector introduce rDNA into host heat-shock electroporation Transformation incubate ligation mixture with ‘competent cells’ cells pretreated to enhance DNA uptake treat according to method 40-41o for 1-2 minutes brief pulse of high voltage

55
Bacterial Transformation with a Plasmid
chromosome E. Coli cell Amps Ampr Permeablize membrane with Ca2+ and heat shock E. Coli cell Ampr + plasmid Select for growth in the presence of ampicillin

57
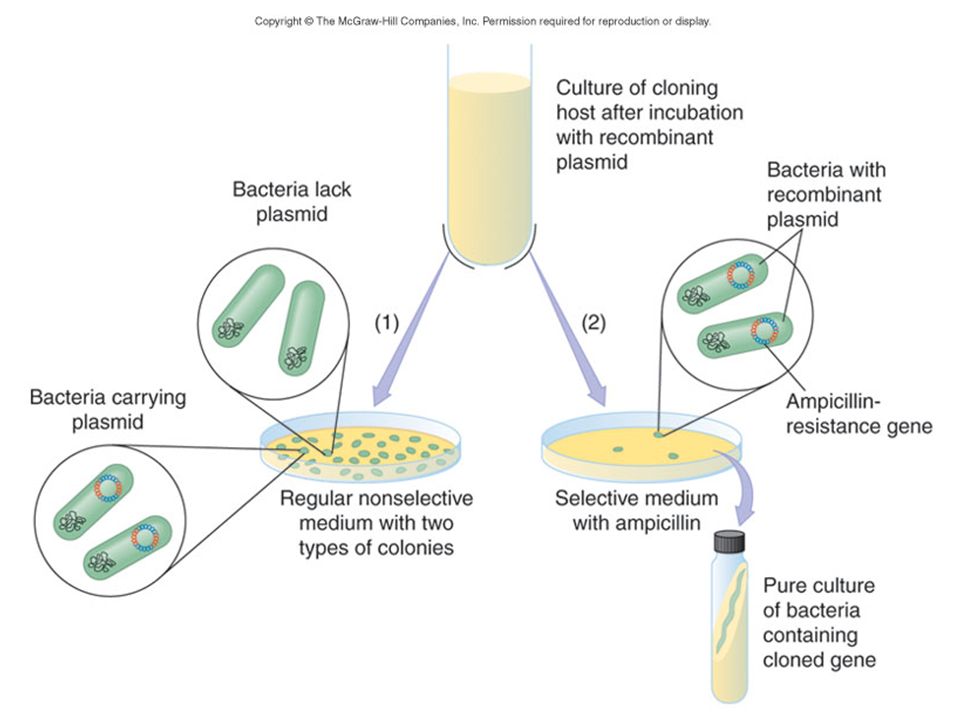
Screening bacteria by replica plating

58
Transformation of E. coli and Selection of Desired Clones
grow E. coli to log phase, optimal for competent cells CaCl2 exposure, maintained at -70° C, then heat shocked at 42 ° C Transformation frequency = transforms / all competent cells Transformation efficiency = transforms / amount of construct DNA E. coli host cell characteristics: No native restriction endonucleases Generally unable to exchange DNA with other E. coli (RecA-) Do not encode for the endA1 endonuclease
 Do not encode for the endA1 endonuclease.")
59
Next Major Advance: Polylinkers
Polylinker is a tandem array of restriction endonuclease sites in a very short expanse of DNA For example, pUC18 polylinker Sites for 13 RE’s Region spans the equivalent of 20 amino acids or 60 nucleotides

60
The Polylinker Advantage
Unique sites (usually) Insert excision facilitated Restriction endonuclease mapping and Subcloning made easier
 Insert excision facilitated. Restriction endonuclease mapping and. Subcloning made easier.")
61
Example 2: pUC19 as a vector used to clone fragments
2,686 bp Ampr gene Regulatable segment of lacZ gene for b-galactosidase EcoRI site not within any coding region Multiple restriction sites for inserting fragments

62
Useful Plasmid Features
Relaxed Replication Selectable Markers Streamlined Polylinker or MCS Identification of Recombinants most derived from pUC or pBR322 |SacI| |ScII| |XbaI||SpeI||BamH||SmaI||PstI||EcRI||EcRV||HIII||ClaI| |SalI||XhoI| |KpnI| GAGCTCCACCGCGGTGGCGGCCGCTCTAGAACTAGTGGATCCCCCGGGCTGCAGGAATTCGATATCAAGCTTATCGATACCGTCGACCTCGAGGGGGGGCCCGGTACC CTCGAGGTGGCGCCACCGCCGGCGAGATCTTGATCACCTAGGGGGCCCGACGTCCTTAAGCTATAGTTCGAATAGCTATGGCAGCTGGAGCTCCCCCCCGGGCCATGG Multiple Cloning Site:

63
a-complementation – relies on modular structure of b-galactosidase
- basic idea is often used with cloning vectors – called insertional inactivation a-complementation LacZ+ - blue colony LacZ- - while colony of you interrupt the lacZ gene, the colony is white Brock Biology of Microorganisms, vol. 9, Chapter 10

64
BamHI digested target DNA
Polylinker from pUC118 EcoR1 SacI KpnI SmaI BamHI XbaI SalI PstI SphI HindIII XmaI AccI HincII Digest with BamHI Add: BamHI digested target DNA Ligated rDNA EcoR1 SacI KpnI SmaI BamHI XmaI BamHI XbaI SalI PstI SphI HindIII AccI HincII

65
Major Advance: Blue/White Screening
E. coli normally produces -galactosidase Production is under control by the lac operon

66
Z = beta galactosidase, Y = lactose permease
Z = beta galactosidase, Y = lactose permease. A = thiogalactoside transactylase, lacI = repressor, Pi = promoter for the lac repressor, P and O = promoter and operator

67
The lac operon with lactose present

68
The lac operon when lactose is absent

69
Major Advance: Blue/White Screening
Host E. coli strains have a deletion in the N-terminus region of the lacZ gene These cloning vectors have the P, O and first 58 amino acids of the lacZ gene (alpha peptide) Intact, the vector in an alpha mutant strain (ex. JM109) produces beta-galactosidase
 Intact, the vector in an alpha mutant strain (ex. JM109) produces beta-galactosidase.")
70
Identifying Recombinants
based on interruption of a gene eg., lacZ gene = b-galactosidase intact b-galactosidase produces blue color in presence of X-gal -complementation or blue-white screening

71
How does a-complementation work?
It all comes down to b-galactosidase Certain strains supply the b-Gal fragment When a is supplied in trans, this allows b-Gal to function Current Protocols in Molecular Biology

72
Which cells have the pUC19-fragment constructs?
IPTG (isopropyl-B-D-tiogactopyranoside) is an inducer of the lac operon regulation Plate the transforms onto ampicillin, IPTG and X-gal plates If no fragment inserted, transform will express b-galactosidase, and it will convert X-gal into a blue product.
 is an inducer of the lac operon regulation. Plate the transforms onto ampicillin, IPTG and X-gal plates. If no fragment inserted, transform will express b-galactosidase, and it will convert X-gal into a blue product.")
73
LB/ampicllin/X-Gal/IPTG plate showing colonies of the
E. coli JM109 competent cells for the white- blue selection

74
Major Advance: Blue/White Screening
When insert DNA in cloned into the polylinker separating the start codon and the alpha peptide sequence in the vector, the peptide is not made Hence, bacterial colonies containing vector plus insert DNA are white

75
Major Advance: Blue/White Screening
Inducer ISOPROPYL-ß-D-THIOGALACTOPYRANOSIDE (IPTG) Colorimetric indicator 5-Bromo-4-chloro-3-indoxyl-beta-D-galactopyranoside (X-gal), colonies turn blue
 Colorimetric indicator 5-Bromo-4-chloro-3-indoxyl-beta-D-galactopyranoside (X-gal), colonies turn blue.")
76
The Major Limitation of Cloning in Plasmids
Upper limit for clone DNA size is 10 kb Requires the preparation of “competent” host cells If the E. coli genome contains 4,639,221 base pairs, how many plasmid clones are needed to clone the entire genome?

77
The Major Limitation of Cloning in Plasmids
Inefficient from generating genomic libraries as overlapping regions needed to place in proper sequence Preference for smaller clones to be transformed Lose large genetic regions

78
Ligation Reaction mix foreign and vector DNA in presence of DNA ligase
optimal ratios of vector to insert generally 1.5-2:1 intermolecular base-pairing can occur between compatible overhangs

79
Intramolecular vs. Intermolecular

80
Kinases and Phosphatases
add or remove phosphate groups and the 5’ ends of DNA or RNA. Kinase +ATP HO-GATC… PO4-GATC… Phosphatase gamma O A (P)-(P)-(P)- alpha beta - the enzyme is not sequence-specific
-(P)-(P)- alpha. beta. - the enzyme is not sequence-specific.")
81
Removal of 5’-PO4 Prevents Vector Self Ligation

83
Purification of Plasmids
Takes advantage of distinct topological state of plasmids. - plasmids will be covalently closed, negatively wound circles when E. coli is lysed. - chormosomal DNA will be sheared into linear, non-topologically constrained fragments (because so big). This difference can be exploited to allow purification of plasmids: - difference in binding ethidium bromide, leading to different densities (CsCl banding, right). - Different rater of re-associate of two strands following denaturation by boiling or alkaline treatment
. This difference can be exploited to allow purification of plasmids: - difference in binding ethidium bromide, leading to different densities (CsCl banding, right). - Different rater of re-associate of two strands following denaturation by boiling or alkaline treatment.")
84
Plasmid conformation affects gel mobility
Figures 4.1 & 4.2

86
Provides extremely tight regulation and extremely strong expression
Invitrogen website The alkaline phosphatase from E. coli was cloned downstream of PBAD

87
Controlled expression systems- many different varieties
araC-PBAD cassette is now marketed by Invitrogen as a controlled expression system 1. Tight repression in the absence of arabinose (and presence of glucose) 2. Strong activation with addition of arabinose Multipartite repression loop Class I transcription activator Schleif, 1996, Chapter 83, Escherichia coli and Salmonella, ASM Press
 2. Strong activation with addition of arabinose. Multipartite repression loop. Class I transcription activator. Schleif, 1996, Chapter 83, Escherichia coli and Salmonella, ASM Press.")
88
Gene fusion systems – monitor the activity of a gene by fusing it to another
Current favorites are the autofluorescent proteins HeLa cells expressing gfp and rfp Clontech website

89
Phages as Vectors Have a natural advantage over plasmids: they infect cells much more efficiently than plasmids transform cells, so the yield of clones with phage vectors is usually higher

90
Cloning in Charon 4 λ DNA At least 12 kb Max. 20 kb Replacement vector

91
Selection of positive genomic clones by Plaque hybridization
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Selection of positive genomic clones by Plaque hybridization

92
Cosmids Cos (cohesive ends), mids=plasmid Accommodate 40-50 kb
Recombinant cosmids are packaged into phage particles. These, cannot replicate as phages but they are infectious so they carry their recombinant DNA into bacterial cells

93
M13 Phage Vectors Single-stranded DNA Site-directed mutagenesis
DNA sequencing

94
Obtaining single-stranded DNA by cloning in M13 phage
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Obtaining single-stranded DNA by cloning in M13 phage

95
Phagemids Characteristics of both phages and plasmids
e.g. pBluescript (pBS)
")
96
pBluescript origin of replication ampicillin resistance gene MCS
A widely used plasmid cloning vector ampicillin resistance gene MCS MCS, Multiple Cloning Site

97
cDNA cloning cDNA is a DNA copy of the mRNA
cDNA library is a set of clones representing as many as possible of the mRNA in a given cell type at a given time

98
Making a cDNA Library Nick translation Fig. 4.14
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Nick translation Fig. 4.14 Making a cDNA Library

99
Copyright © The McGraw-Hill Companies, Inc
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display.

100
Using RT-PCR in cDNA Cloning

101
Using RT-PCR to clone a single cDNA whose sequence is known
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. Using RT-PCR to clone a single cDNA whose sequence is known

102
What kind of vector should be used to ligate a cDNA?
A plasmid or phagemid vector such as pUC or pBS can be used; if so, positive clones are usually identified by colony hybridization with a labeled DNA probe Or a λ vector, such as λgt11, as vector → places the cloned cDNA under the control of lac promoter so that transcription and translation can occur One can use an antibody to screen for the product

103
Rapid Amplification of cDNA Ends (RACE)
")
104
5' RACE to fill in the 5' end of a cDNA
Copyright © The McGraw-Hill Companies, Inc. Permission required for reproduction or display. 5' RACE to fill in the 5' end of a cDNA

105
One major issue is the size of the insert DNA
The larger the fragment, the more of the source genome can be represented on one plasmid For a gene library, we can easily calculate the number of individual recombinant molecules required to represent an entire genome N = [ln (1-P)]/[ln (1-f)] For the human genome and a standard plasmid P – 99% or 0.99 confidence f – 5 kb/4,400,000 kb = 1.14 x 10-6 P – probability of complete coverage N – number of individual clones required f – proportion of genome in average fragment N = [ln .01]/[ln ] = -4.6/-1.14 x 10-7 = 40,350,877 individuals Plasmid vectors stable inserts of 5 kb
]/[ln (1-f)] For the human genome and a standard plasmid. P – 99% or 0.99 confidence. f – 5 kb/4,400,000 kb = 1.14 x P – probability of complete coverage. N – number of individual clones required. f – proportion of genome in average fragment. N = [ln .01]/[ln ] = -4.6/-1.14 x = 40,350,877 individuals. Plasmid vectors. stable inserts of 5 kb.")
106
Bacteriophage vectors can acommodate larger inserts
larger inserts than plasmids – introduced by phage infection of cells 20 – 35 kb inserts For the human genome and a standard plasmid P – 99% or 0.99 confidence f – 35 kb/4,400,000 kb = 7.95 x 10-6 N = [ln .01]/[ln ] = -4.6/-7.95 x 10-6 = 578, 616 individuals A big improvement!

107
A even bigger improvement!
Yeast artificial chromsomes (YACs) and bacterial artificial chromsomes (BACs) Can accomodate from 300 – 500 kb of DNA – great for large genomes For the human genome and a standard plasmid P – 99% or 0.99 confidence f – 500 kb/4,400,000 kb = 1.14 x 10-4 N = [ln .01]/[ln ] = -4.6/-1.14 x 10-4 = 40,350 individuals A even bigger improvement!
 and bacterial artificial chromsomes (BACs) Can accomodate from 300 – 500 kb of DNA – great for large genomes. For the human genome and a standard plasmid. P – 99% or 0.99 confidence. f – 500 kb/4,400,000 kb = 1.14 x N = [ln .01]/[ln ] = -4.6/-1.14 x = 40,350 individuals. A even bigger improvement!")
108
Whats a YAC? Yeast artificial chromsome
self-replicating vector that can be maintained in yeast Can accommodate large insert fragments Reeves et al., 1992, Methods Enzymol. 216:

109
What are BACs? bacterial artificial chromsomes
Derived from the F plasmid of E. coli - very stably maintained - 1-2 copies per cell (strict copy number contro) What are BACs? bacterial artificial chromsomes - very low copy number vectors that can accomodate huge inserts Shizuya et al, 1992, PNAS 89:
 What are BACs bacterial artificial chromsomes. - very low copy number vectors that can accomodate huge inserts. Shizuya et al, 1992, PNAS 89:")
Podobne prezentacje






klasyczne>")












