Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
BIOLOGIA KOMÓRKI (WYKŁAD 1, cz
BIOLOGIA KOMÓRKI (WYKŁAD 1, cz.2, BIOLOGIA Z GENETYKĄ, KIERUNEK FIZYKA MEDYCZNA) Cykl komórkowy Podział komórki somatycznej - MITOZA Fazy cyklu komórkowego (G1, S, G2, M) Regulacja cyklu komórkowego (cykliny, kinazy zależne od cyklin, inhibitory i promotory przejścia z fazy do fazy) Skutki zaburzenia cyklu komórkowego (śmierć komórki, transformacja nowotworowa) Szlaki przekazywania sygnałów Receptory powierzchniowe i ligandy Receptory wewnątrzkomórkowe i ligandy Nekroza (czynniki pobudzające nekrozę) Apoptoza (szlaki apoptotyczne, czynniki blokujące i pobudzające apoptozę Aleksander L. Sieroń
 Cykl komórkowy. Podział komórki somatycznej - MITOZA. Fazy cyklu komórkowego (G1, S, G2, M) Regulacja cyklu komórkowego (cykliny, kinazy zależne od cyklin, inhibitory i promotory przejścia z fazy do fazy) Skutki zaburzenia cyklu komórkowego (śmierć komórki, transformacja nowotworowa) Szlaki przekazywania sygnałów. Receptory powierzchniowe i ligandy. Receptory wewnątrzkomórkowe i ligandy. Nekroza (czynniki pobudzające nekrozę) Apoptoza (szlaki apoptotyczne, czynniki blokujące i pobudzające apoptozę. Aleksander L. Sieroń.")
2
BIOLOGIA KOMÓRKI (WYKŁAD 1, cz
BIOLOGIA KOMÓRKI (WYKŁAD 1, cz.2, BIOLOGIA Z GENETYKĄ, KIERUNEK FIZYKA MEDYCZNA) Platforma e-learning Klucz embrio318 Aleksander L. Sieroń
 Platforma e-learning. Klucz embrio318. Aleksander L. Sieroń.")
3
WYKŁAD 2 (BIOLOGIA MEDYCZNA; KIERUNEK LEKARSKI)
CYKL KOMÓRKOWY FAZY CYKLU REGULATORY CYKLU AKTYWATORY BLOKERY APOPTOZA SZLAKI MEJOZA Aleksander L. Sieroń

4
Cykl komórkowy 0,5 DO 24 GODZIN Bloker guzów geny, CDK Blokowanie
Czynniki wzrostu Oncogeny Cykliny & CDK Bloker guzów geny, CDK Blokowanie „Punkt zakazu” (Niemożliwość powrotu) Nowa komórka siostrzana Mitoza (podział komórek) Początek cyklu 25% 10% 25% Cykl komórkowy 0,5 DO 24 GODZIN Synteza, (Podwojenie DNA) 40% Aleksander L. Sieroń
 Nowa komórka siostrzana. Mitoza (podział komórek) Początek. cyklu. 25% 10% 25% Cykl komórkowy. 0,5 DO 24 GODZIN. Synteza, (Podwojenie. DNA) 40% Aleksander L. Sieroń.")
5
Cykl komórkowy 0,5 DO 24 GODZIN Nowa komórka siostrzana
Mitoza (podział komórek) Początek cyklu Czynniki wzrostu Onkogeny Cykliny & CDK Bloker guzów geny, CDK Blokowanie Synteza, (Podwojenie DNA) „Punkt zakazu” (Niemożliwość powrotu) Cykl komórkowy 0,5 DO 24 GODZIN Aleksander L. Sieroń
 Początek. cyklu. Czynniki wzrostu. Onkogeny. Cykliny & CDK. Bloker guzów. geny, CDK. Blokowanie. Synteza, (Podwojenie. DNA) „Punkt zakazu (Niemożliwość powrotu) Cykl komórkowy. 0,5 DO 24 GODZIN. Aleksander L. Sieroń.")
6
Segregacja chromosomów
CYKL KOMÓRKOWY Segregacja chromosomów Podział komórki 2 Wielkość Komórki 1 2 (4C) Zawartość DNA 1 (2C) Replikacja DNA CDK1 activity Cyclin levels Aleksander L. Sieroń
 Zawartość. DNA. 1 (2C) Replikacja DNA. CDK1 activity. Cyclin levels. Aleksander L. Sieroń.")
7
WYKŁAD 2 (BIOLOGIA Z GENETYKĄ; KIERUNEK FIZYKA MEDYCZNA)
CYKL KOMÓRKOWY FAZY CYKLU REGULATORY CYKLU AKTYWATORY BLOKERY APOPTOZA SZLAKI MEJOZA Aleksander L. Sieroń

8
Segregacja chromosomów (kinaz cyklino-zależnych)
CYKL KOMÓRKOWY Segregacja chromosomów Podział komórki 2 Wielkość komórki 1 2 Zawartość DNA 1 Replikacja DNA Stężenie Cyklin Aktywność CDK (kinaz cyklino-zależnych) Aleksander L. Sieroń
 Aleksander L. Sieroń.")
9
Cykl komórkowy jest regulowany przez CDK (kinazy zależne od cyklin).
POCZĄTEK CYKLU

10
To Cykliny aktywują CDK
POCZĄTEK CYKLU

11
Niskocząsteczkowe białka, p9cks i p15cdk-BP,
ściśle wiążą się z CDK modulując ich aktywność POCZĄTEK CYKLU

12
Aktywatory kompleksów cyklina/CDK:
fosfatazy białkowe (cdc25) i inne kinazy (cdk7/cyklina H, polo) również regulują aktywność kompleksów cykliny/CDK. POCZĄTEK CYKLU
 i inne kinazy (cdk7/cyklina H, polo) również regulują aktywność kompleksów cykliny/CDK. POCZĄTEK CYKLU.")
13
Strukturalny mechanizm
aktywacji Cdk Chapter 3 The Cell-Cycle Control System© 2007 New Science Press Ltd

14
WYKŁAD 2 (BIOLOGIA MEDYCZNA; KIERUNEK LEKARSKI)
CYKL KOMÓRKOWY FAZY CYKLU REGULATORY CYKLU AKTYWATORY BLOKERY APOPTOZA SZLAKI MEJOZA Aleksander L. Sieroń

15
Blokery aktywności CDK:
Liczne białka blokują aktywność CDK wiążąc się z nimi, stechiometrycznie (CIP1, INK4) lub przez modyfikacje enzymatyczne (wee1, KAP, etc.) POCZĄTEK CYKLU BRCA1
 lub. przez modyfikacje enzymatyczne (wee1, KAP, etc.) POCZĄTEK CYKLU. BRCA1.")
16
liczne enzymy utrzymują w G2 nieaktywny kompleks cdk1/cyklina B
Przejście do mitozy: liczne enzymy utrzymują w G2 nieaktywny kompleks cdk1/cyklina B nieaktywny aktywny G2 MITOZA (M) nieaktywny aktywny PROFAZA MITOZA nieaktywny aktywny PROFAZA MITOZA
 nieaktywny. aktywny. PROFAZA. MITOZA. nieaktywny. aktywny. PROFAZA. MITOZA.")
17
G2 MITOZA (M) nieaktywny aktywny Przejście profaza/metafaza:
Uszkodzenie DNA zatrzymuje komórki w G2 przez wpływ na aktywność cdc25 nieaktywny aktywny MITOZA (M) G2
 G2.")
18
Przejście profaza/metafaza: Inne czynniki regulatorowe!
MITOZA (M)
")
19
Oznaczanie aktywności kinazowej CDK1/cyklina B
Histon H1 +32P-ATP 32P-Histon H1 + ADP Pomiar aktywności kinazowej cdk1/cyklina B

20
Potencjalne miejsca blokowania aktywności CDK
Miejsca fosforylowane Miejsca oddziaływania CDC25 Miejsca wiązania blokerów białkowych Kieszeń wiązania ATP Miejsca wiązania substratu Pętla fosfotreoniny Domena wiązania CKS CYKLIN „Skrzynka rozpadowa” Komórkowa lokalizacja domen

21
Partnerzy CDK

22
Zatrzymanie cyklu komórkowego
Onkogeny (np. E1A, Myc) Uszkodzenie DNA (UV, leki) p19ARF Kinazy (np. ATM, DNAPK) p53 Zatrzymanie cyklu komórkowego lub apoptoza Aleksander L. Sieroń
 Uszkodzenie DNA. (UV, leki) p19ARF. Kinazy. (np. ATM, DNAPK) p53. Zatrzymanie cyklu komórkowego. lub apoptoza. Aleksander L. Sieroń.")
23
(UV, PROMIENIOWANIE JONIZUJĄCE, LEKI, ITP.)
USZKODZENIE DNA (UV, PROMIENIOWANIE JONIZUJĄCE, LEKI, ITP.) Blok MDM2 stabilizacja p53 Blok MDM4 p53 p21 *CYKLINA E/CDK2 CYKLINA-CDK ATP ADP *E2F CYKLINA + CDK Rb:E2F ppRb ARESZT W G1 S A.L. SIEROŃ; 2005/06
 Blok MDM2. stabilizacja p53. Blok. MDM4. p53. p21. *CYKLINA E/CDK2. CYKLINA-CDK. ATP. ADP. *E2F. CYKLINA + CDK. Rb:E2F. ppRb. ARESZT W. G1. S. A.L. SIEROŃ; 2005/06.")
24
(ataxia telangiectasia mutated/ATM and Rad3-related)
USZKODZENIA W KOMÓRCE ATM/ATR (ataxia telangiectasia mutated/ATM and Rad3-related) ATM, ATR i hCds1/Chk2 są białkami odpowiadającymi na uszkodzenia w komórce zmieniając fosforylację produktu genu BRCA ? hCds1/Chk2 BRCA1 Chk1 – Kinaza regulatorowa Cdc25C Kinaza Wee1 Cyklina B/Cdk1 G2 M ARESZT W Aleksander L. Sieroń
 ATM, ATR i hCds1/Chk2. są białkami. odpowiadającymi na. uszkodzenia w komórce. zmieniając fosforylację. produktu genu BRCA. hCds1/Chk2. BRCA1. Chk1 – Kinaza regulatorowa. Cdc25C. Kinaza. Wee1. Cyklina B/Cdk1. G2. M. ARESZT W. Aleksander L. Sieroń.")
25
Cykl komórkowy cdk 1 cykliny A i B pRB/RIZ1 p53 p21 pRB cdk2, 4 i 6
Nowa komórka siostrzana Mitoza (podział komórek) Początek cyklu Czynniki wzrostu Oncogeny Cykliny & CDK Bloker guzów geny, CDK Blokowanie Synteza, (Podwojenie DNA) „Punkt zakazu” (Niemożliwość powrotu) Cykl komórkowy cdk 1 cykliny A i B pRB/RIZ1 Wejście do Apoptozy p53 p21 pRB cdk2, 4 i 6 cykliny A, E i D Wyjście do G0 Wejście do Apoptozy Aleksander L. Sieroń
 Początek. cyklu. Czynniki wzrostu. Oncogeny. Cykliny & CDK. Bloker guzów. geny, CDK. Blokowanie. Synteza, (Podwojenie. DNA) „Punkt zakazu (Niemożliwość powrotu) Cykl komórkowy. cdk 1. cykliny A i B. pRB/RIZ1. Wejście. do. Apoptozy. p53. p21. pRB. cdk2, 4 i 6. cykliny A, E i D. Wyjście. do G0. Wejście. do. Apoptozy. Aleksander L. Sieroń.")
26
WYKŁAD 2 (BIOLOGIA MEDYCZNA; KIERUNEK LEKARSKI)
CYKL KOMÓRKOWY FAZY CYKLU REGULATORY CYKLU AKTYWATORY BLOKERY APOPTOZA SZLAKI MEJOZA Aleksander L. Sieroń

27
ŚMIERĆ KOMÓRKI I APOPTOZA
Śmierć komórek następuje przez jeden z dwóch mechanizmów: 1. NEKROZĘ - następującą w wyniku działania uszkadzających czynników zewnętrznych np.: ciężkie oparzenia urazy mechaniczne 2. APOPTOZĘ - czyli zaprogramowaną śmierć komórek - będącą wynikiem następujących kolejno po sobie reakcji na bodźce biochemiczne lub fizyczne. Aleksander L. Sieroń

28
APOPTOZA Jest złożonym sposobem pozbywania się przez organizm niechcianych lub uszkodzonych komórek. Jest zaangażowana w homeostazę tkanek i różnicowanie komórek. Większość komórek organizmu ma z góry określoną długość życia. Śmierć komórki jest normalnie, ściśle regulowanym procesem, w którym komórki nieustannie odpowiadają na sygnały chemiczne pochodzące od innych komórek lub ze swojego otoczenia. Aleksander L. Sieroń

29
APOPTOZA jest bezpośrednio zaangażowana
w degeneracji; np. choroby: Alzheimera, Huntigtona, Parkinsonizm. w autoimmunoagresji: odczyny reumatyczne w choroby wirusowe: AIDS w przemianach nowotworowych Aleksander L. Sieroń

30
S T A R T Apoptoza zaczyna się w komórkach nie dających się naprawić, lub które zakończyły swoją zaprogramowaną funkcję biologiczną. Inna morfologia komórek wynika z zaburzeń w budowie błon komórkowych. Aleksander L. Sieroń

31
WYKŁAD 2 (BIOLOGIA MEDYCZNA; KIERUNEK LEKARSKI)
CYKL KOMÓRKOWY FAZY CYKLU REGULATORY CYKLU AKTYWATORY BLOKERY APOPTOZA SZLAKI MEJOZA Aleksander L. Sieroń

32
DOMENA ZEWNĄTRZ KOMÓRKOWA
Complex DOMENA BŁONOWA DOMENA ZEWNĄTRZ KOMÓRKOWA CYTOPLAZMATYCZNA DOMENA

33
Complex ZEWNĄTRZ CYTOPLAZMA

34
A P O P T O Z A SZLAKI APOPTOZY RECEPTOROWY MITOCHONDRIALNY
A.L. SIEROŃ; 2005/06

35
Szlak mitochondrialny
Szlak receptorowy FasL Domena zewnątrz-komórkowa Fas/CD95 Błona komórkowa Szlak mitochondrialny pro--Bid cytochrom c kaspaz-9/cytochrom c Kaspazy wykonawcze & inne substraty mitochondria p15tBid FADD pro-kaspaza-8 Aleksander L. Sieroń

36
ODCZYNY ZAPALNE A P O P T O Z A
(Cys-Proteazy) ODCZYNY ZAPALNE A P O P T O Z A
 ODCZYNY ZAPALNE. A P O P T O Z A.")
37
Po przyłączeniu ligandu (np..: Fas)
Szlak receptorowy Domena zewnątrz-komórkowa FasL Fas/CD95 Błona komórkowa Po przyłączeniu ligandu (np..: Fas) Szlak mitochondrialny FADD cytochrom c kaspaza-9/cytochrom c Kaspazy wykonawcze & inne substraty mitochondria p15tBid pro--Bid pro-kaspaza-8 pro-kaspaza-8 kaspaza 8 Aleksander L. Sieroń
 Szlak mitochondrialny. FADD. cytochrom c. kaspaza-9/cytochrom c. Kaspazy wykonawcze & inne substraty. mitochondria. p15tBid. pro--Bid. pro-kaspaza-8. pro-kaspaza-8. kaspaza 8. Aleksander L. Sieroń.")
38
Szlak mitochondrialny
Szlak receptorowy pro-kaspaza-8 pro--Bid kaspaza 8 cytochrom c kaspaza-9/cytochrom c mitochondria p15tBid Domena zewnątrz-komórkowa FasL Fas/CD95 Błona komórkowa FADD Szlak mitochondrialny kaspaza 3 Kaspazy wykonawcze & inne substraty Aleksander L. Sieroń

39
MUTACJE NIEAKTYWNE LIGANDY
pro-kaspaza-8 Domena zewnątrz-komórkowa FasL Fas/CD95 Błona komórkowa FADD Jądro komórkowe Bcl2 MUTACJE NIEAKTYWNA KASPAZA 8 MUTACJE NIEAKTYWNE RECEPTORY p53 Bax MUTACJE NIEAKTYWNE KINAZY Kaspazy wykonawcze & inne substraty kaspaza 3 cytochrom c kaspaz-9/cytochrom c p15tBid mitochondria Aleksander L. Sieroń

40
? Uszkodzenie DNA Zanik Telomerów Onkogeny
Model apoptotycznej odpowiedzi komórek na cytotoksyny, napromieniowanie i niedobór cytokin. Białka BH3-tylko Puma, Noxa i Bim pośredniczą, zarówno w zależnej, jak i niezależnej od p53 odpowiedzi na stres. Odpowiedzi te są aktywowane jednakowo przez stres wymuszony (niedobór cytokin, cytotoksyny i promieniowanie), jak i sygnały o stresie, takie które powstają w czasie tumorogenezy od aktywowanych onkogenów, zaniku telomerów i w hypoksji. Aktywacja kinazy ATM lub białka Arf blokuje białko Mdm2, co wywołuje wzrost stężenia p53 w komórkach. P53 indukuje transkrypcję genów p21, Puma i Noxa, a także innych o słabo poznanym znaczeniu (pola zakreskowane). Aktywowanie Bcl-2 (i innych pokrewnych mu białek) przez białka BH3-tylko powoduje aktywację Bax i Bak co wzbudza apoptozę. Skróty: STS - staurosporyna; dex - deksametazon. ATM Arf ? Mdm2 ? Hypoksja MDM4 i 2 p53 Niedobór cytokin Wypływ Ca2+ p21 Taksol PMA Zatrzymanie Cyklu komórkowego STS dex PIG3, PIG8, PERP P53AIP1, p53DINP1 Bim Puma Noxa ? Bcl-2 Bax/ Bak Apoptoza Villunger A. et al. (2003) Science, 302:
, jak i sygnały o stresie, takie które powstają w czasie tumorogenezy od aktywowanych onkogenów, zaniku telomerów i w hypoksji. Aktywacja kinazy ATM lub białka Arf blokuje białko Mdm2, co wywołuje wzrost stężenia p53 w komórkach. P53 indukuje transkrypcję genów p21, Puma i Noxa, a także innych o słabo poznanym znaczeniu (pola zakreskowane). Aktywowanie Bcl-2 (i innych pokrewnych mu białek) przez białka BH3-tylko powoduje aktywację Bax i Bak co wzbudza apoptozę. Skróty: STS - staurosporyna; dex - deksametazon. ATM. Arf. Mdm2. Hypoksja. MDM4 i 2. p53. Niedobór. cytokin. Wypływ Ca2+ p21. Taksol. PMA. Zatrzymanie. Cyklu komórkowego. STS. dex. PIG3, PIG8, PERP. P53AIP1, p53DINP1. Bim. Puma. Noxa. Bcl-2. Bax/ Bak. Apoptoza. Villunger A. et al. (2003) Science, 302:")
41
Rycina 6 Model wpływu Bcl-xS w fibroblastach zarodków mysich (MEFs).
Lindenboim L., et al. Cell Death and Differentiation (2005) 12, 713–723 Rycina 6 Model wpływu Bcl-xS w fibroblastach zarodków mysich (MEFs). Ekspresja Bcl-xS w MEFs eksponuje N-koniec (NT) Bak, co prowadzi do aktywacji Bak. Zaktywowany Bak może indukować aż trzy ścieżki sygnałowe: główna ścieżka prowadzi do uwolnienia cytochromu c, który aktywuje apoptosom i nastepczą śmierć komórki na drodze zależnej od kaspaz; druga ścieżka, prowadzi do Apaf-1- i niezależnej od kaspazy-9 śmierci komórki; a trzecia ścieżka wzbudza ekspozycję NT Nax aktywując go na obydwu drogach zależnej- i niezależnej od caspazy-9. Pierwsze dwie ścieżki mają udział w procesach śmierci. Znaczenie trzeciej ścieżki jest dotychczas słabo poznane.
 12, 713–723. Rycina 6 Model wpływu Bcl-xS w fibroblastach zarodków mysich (MEFs). Ekspresja Bcl-xS w MEFs eksponuje N-koniec (NT) Bak, co prowadzi do aktywacji Bak. Zaktywowany Bak może indukować aż trzy ścieżki sygnałowe: główna ścieżka prowadzi do uwolnienia cytochromu c, który aktywuje apoptosom i nastepczą śmierć komórki na drodze zależnej od kaspaz; druga ścieżka, prowadzi do Apaf-1- i niezależnej od kaspazy-9 śmierci komórki; a trzecia ścieżka wzbudza ekspozycję NT Nax aktywując go na obydwu drogach zależnej- i niezależnej od caspazy-9. Pierwsze dwie ścieżki mają udział w procesach śmierci. Znaczenie trzeciej ścieżki jest dotychczas słabo poznane.")
42
PRZEBIEG APOPTOZY W jądrze komórkowym („mózgu” komórki) chromatyna ulega zagęsz-czeniu, a DNA fragmentacji. Komórka w apoptozie jest otaczana przez sąsiadujące normalne komór-ki, które pochłaniają jej fragmenty i zużywają je na własne potrzeby. Aleksander L. Sieroń

43
Główne ścieżki prowadzące do śmierci kaspozo -zależnej i –niezależnej
Główne ścieżki prowadzące do śmierci kaspozo -zależnej i –niezależnej. Zidentyfikowano dwie ścieżki apoptotyczne kaspazo-zależne: ścieżkę zewnętrzną pobudzaną przez czynniki należące do nadrodziny typu receptora TNF-NGF (czynnik wzrostu nerwu), takie jak receptor TNF (TNFR), CD95 (Fas)-APO-1 receptor lub receptor typu TRAIL (‘receptory śmierci’) oraz ścieżkę wewnętrzną z udziałem MOMP prowadzącym do formowania kompleksów aktywujących kaspazy pomiędzy kaspazą 9 i Apaf-1 (apoptosom). Pobudzenie receptora śmierci prowadzi zwykle do rekrutacji i aktywacji kaspazy 8 za pośrednictwem białek adaptorowych FADD i TRADD, tworzących DISC, który rozprzestrzenia sygnał śmierci dwiema drogami: przez proteolizę białka BH3-only protein Bid, co wywołuje przemieszczenie tego ostatniego do mitochondrium i MOMP oraz przez bezpośrednią proteolizę następnych kaspaz, która powoduje ich aktywację. Na ścieżce wewnętrznej białka BH3-only działają tylko w odpowiedzi na stres komórkowy, uszkodzenie lub infekcję i mogą być mobilizowane do pobudzania MOMP w drodze modyfikacji potranslacyjnych. Białka BH3-only najprawdopodobniej wzbudzają MOMP poprzez zapoczątkowanie oligomeryzacji Bax i/lub Bak w zewnętrznej błonie mitochondrialnej, w której tworzą kanały, przez które uciekają liczne białka z przestrzeni międzybłonowej. W odniesieniu do uszkodzenia DNA stabilizacja białka supresora guzów p53 może prowadzić do aktywacji transkrypcji białek BH3-only, Puma i Noxa promujących MOMP poprzez kanał Bax-Bak. Alternatywna ścieżka apoptozy zależnej od p53 proponuje mechanizm z udziałem transkrypcyjnej nadregulacji białka PIDD. PIDD może promować tworzenie kompleksu własnego z RAIDD i kaspazą 2 (‘piddosomu’). Nie wiadomo dokładnie, jak piddosom może promować śmierć komórki, ale może w tym procesie uczestniczyć MOMP zależny od kaspazy 2. Niektóre białka mitochondrialne uwolnione przez MOMP (AIF, HtrA2/Omi, endonukleaza G) mogą promować śmierć komórki niezależną od kaspazy poprzez jeszcze bardzo słabo poznany mechanizm. Śmierć komórki niezależna od kaspaz może być także wynikiem pobudzeń prowadzących do zwiększonej przepuszczalności błony lizosomalnej (LMP) i zwiększonego uwalniania proteaz katepsynowych.
, takie jak receptor TNF (TNFR), CD95 (Fas)-APO-1 receptor lub receptor typu TRAIL (‘receptory śmierci’) oraz ścieżkę wewnętrzną z udziałem MOMP prowadzącym do formowania kompleksów aktywujących kaspazy pomiędzy kaspazą 9 i Apaf-1 (apoptosom). Pobudzenie receptora śmierci prowadzi zwykle do rekrutacji i aktywacji kaspazy 8 za pośrednictwem białek adaptorowych FADD i TRADD, tworzących DISC, który rozprzestrzenia sygnał śmierci dwiema drogami: przez proteolizę białka BH3-only protein Bid, co wywołuje przemieszczenie tego ostatniego do mitochondrium i MOMP oraz przez bezpośrednią proteolizę następnych kaspaz, która powoduje ich aktywację. Na ścieżce wewnętrznej białka BH3-only działają tylko w odpowiedzi na stres komórkowy, uszkodzenie lub infekcję i mogą być mobilizowane do pobudzania MOMP w drodze modyfikacji potranslacyjnych. Białka BH3-only najprawdopodobniej wzbudzają MOMP poprzez zapoczątkowanie oligomeryzacji Bax i/lub Bak w zewnętrznej błonie mitochondrialnej, w której tworzą kanały, przez które uciekają liczne białka z przestrzeni międzybłonowej. W odniesieniu do uszkodzenia DNA stabilizacja białka supresora guzów p53 może prowadzić do aktywacji transkrypcji białek BH3-only, Puma i Noxa promujących MOMP poprzez kanał Bax-Bak. Alternatywna ścieżka apoptozy zależnej od p53 proponuje mechanizm z udziałem transkrypcyjnej nadregulacji białka PIDD. PIDD może promować tworzenie kompleksu własnego z RAIDD i kaspazą 2 (‘piddosomu’). Nie wiadomo dokładnie, jak piddosom może promować śmierć komórki, ale może w tym procesie uczestniczyć MOMP zależny od kaspazy 2. Niektóre białka mitochondrialne uwolnione przez MOMP (AIF, HtrA2/Omi, endonukleaza G) mogą promować śmierć komórki niezależną od kaspazy poprzez jeszcze bardzo słabo poznany mechanizm. Śmierć komórki niezależna od kaspaz może być także wynikiem pobudzeń prowadzących do zwiększonej przepuszczalności błony lizosomalnej (LMP) i zwiększonego uwalniania proteaz katepsynowych.")
44
Shin S. et al. EMBO J. (2005) 24, 3532–3542 Proponowany mechanizm regulacji apoptozy z udziałem TRAIL. (Lewy schemat) Niska aktywność wewnątrzkomórkowego PKCK2 (1) lub wysoka aktywność wewnątrzkomórkowego PKCK2 jest obniżana przez specyficzny inhibitor (1’), defosforylowane monomery prokaspazy-2. Prokaspaza-2 jest następnie aktywowana w wyniku jej dimeryzacji (2), a zaktywowana kaspaza-2 tnie monomer prokaspazy-8 pomiędzy większą i mniejszą podjednostką (3). W takiej sytuacji dochodzi do ‘wzbudzenia’ apoptozy zależnej od TRAIL w komórkach nowotoworowych. Jeżeli TRAIL nie jest połączony z receptorami TRAIL-śmierć, cięta prokaspaza-8 jest kierowana do proteasomu celem degradacji (4). W przypadku przeciwnym TRAIL jest wiązany do swego receptora, przecięta prokaspaza-8 jest rekrutowana przez receptory śmierci TRAIL, czego wynikiem jest utworzenie DISC (4’). Drugie cięcie między prodomeną i podjednostką większą może być wykonane efektywnie w wyniku dimeryzacji ciętych prokaspaz-8, która zachodzi z udziałem DISC (5), co prowadzi do aktywacji prokaspazy-8 (6), a następnie apoptozy za pośrednictwem TRAIL (7). (Prawy schemat) Gdy wewnątrzkomórkowa aktywność PKCK2 jest wysoka prokaspaza-2 nie może być aktywowana; a zatem prokaspaza-8 nie może być przekształcana. Nawet z zaangażowaniem TRAIL, prokaspaza-8 w DISC nie może być aktywowana w pełni i w związku z tym nie wystąpi apoptoza z udziałem TRAIL.
 Niska aktywność wewnątrzkomórkowego PKCK2 (1) lub wysoka aktywność wewnątrzkomórkowego PKCK2 jest obniżana przez specyficzny inhibitor (1’), defosforylowane monomery prokaspazy-2. Prokaspaza-2 jest następnie aktywowana w wyniku jej dimeryzacji (2), a zaktywowana kaspaza-2 tnie monomer prokaspazy-8 pomiędzy większą i mniejszą podjednostką (3). W takiej sytuacji dochodzi do ‘wzbudzenia’ apoptozy zależnej od TRAIL w komórkach nowotoworowych. Jeżeli TRAIL nie jest połączony z receptorami TRAIL-śmierć, cięta prokaspaza-8 jest kierowana do proteasomu celem degradacji (4). W przypadku przeciwnym TRAIL jest wiązany do swego receptora, przecięta prokaspaza-8 jest rekrutowana przez receptory śmierci TRAIL, czego wynikiem jest utworzenie DISC (4’). Drugie cięcie między prodomeną i podjednostką większą może być wykonane efektywnie w wyniku dimeryzacji ciętych prokaspaz-8, która zachodzi z udziałem DISC (5), co prowadzi do aktywacji prokaspazy-8 (6), a następnie apoptozy za pośrednictwem TRAIL (7). (Prawy schemat) Gdy wewnątrzkomórkowa aktywność PKCK2 jest wysoka prokaspaza-2 nie może być aktywowana; a zatem prokaspaza-8 nie może być przekształcana. Nawet z zaangażowaniem TRAIL, prokaspaza-8 w DISC nie może być aktywowana w pełni i w związku z tym nie wystąpi apoptoza z udziałem TRAIL.")
45
Śmierć kaspazo-zależna Śmierć kaspazo-niezależna
Śmierć z aktywacją kaspaz Śmierć z aktywacją kaspaz Śmierć bez aktywacji kaspaz Przeżycie Apoptoza Nekroza Śmierć apoptozo-podobna Śmierć autofagowa Apoptyczna śmierć komórki Niapoptyczna śmierć komórki Wpływ blokerów kaspaz Wyznaczone ścieżki Związki między kaspazo-zależnością i morfotypem śmierci komórki

46
WYKŁAD 2 (BIOLOGIA I BIOLOGIA Z GENETYKĄ; KIERUNEK LEKARSKI)
CYKL KOMÓRKOWY FAZY CYKLU REGULATORY CYKLU AKTYWATORY BLOKERY APOPTOZA SZLAKI MEJOZA Aleksander L. Sieroń

47
MEJOZA – Profaza

48
chromatyda centromer MEJOZA Budowa chromosomu
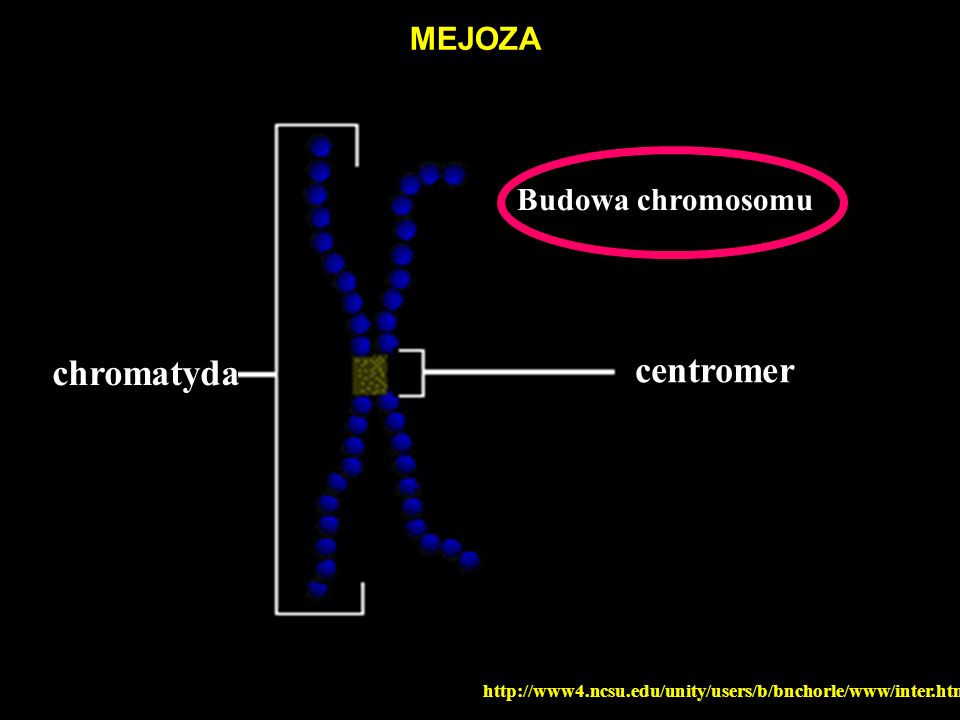
49
MJOZA – Profaza

50
Budowa tetrady (Biwalentu)
chromatydy siostrzane Budowa tetrady (Biwalentu) chromatydy niesiostrzane Liczba Chromosomów = 2n Ilość DNA = 4C
 chromatydy niesiostrzane. Liczba Chromosomów = 2n. Ilość DNA = 4C.")
51
Metafaza I Liczba Chromosomów = 2n Ilość DNA = 4C Włókno wrzeciona
Płytka metacentryczna Liczba Chromosomów = 2n Ilość DNA = 4C

52
Anafaza I Liczba Chromosomów = 2n Ilość DNA = 4C
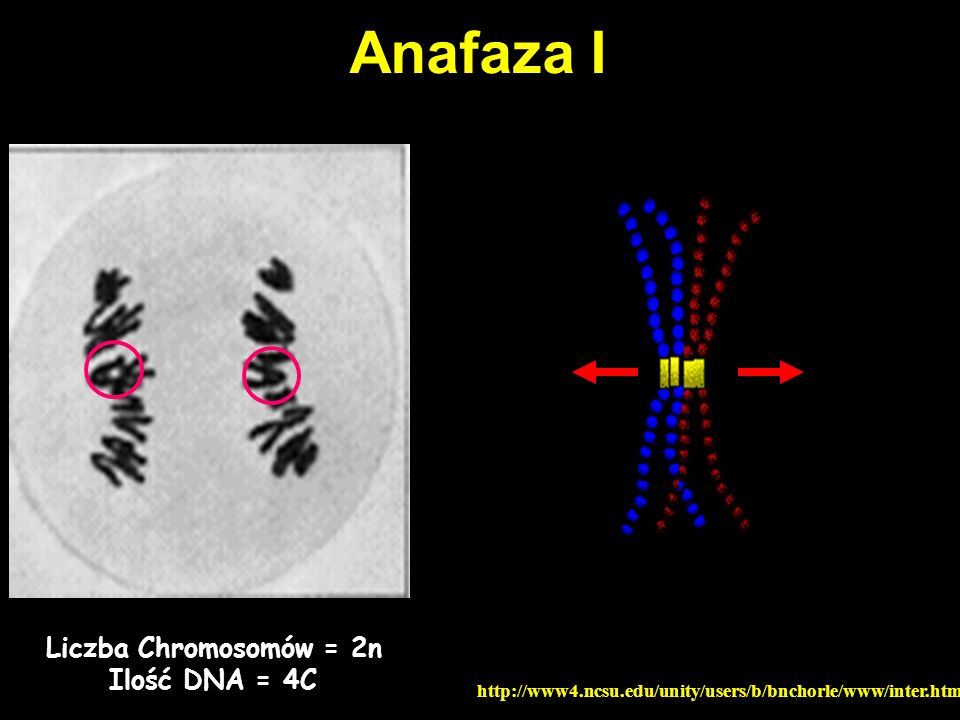
53
PAMIĘTAĆ – 1-szy podział mejotyczny
Telofaza I PAMIĘTAĆ – 1-szy podział mejotyczny to podział REDYKCYJNY Liczba Chromosomów = 1n Ilość DNA = 2C

54
Profaza II Liczba Chromosomów = 1n Ilość DNA = 2C
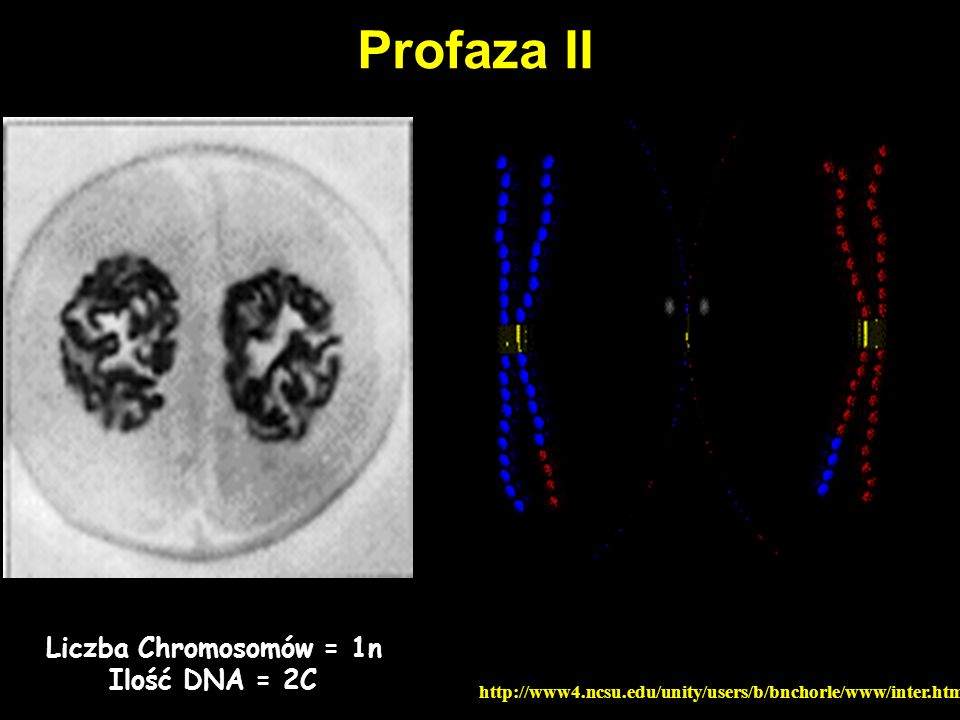
55
Metafaza II Liczba Chromosomów = 1n Ilość DNA = 2C
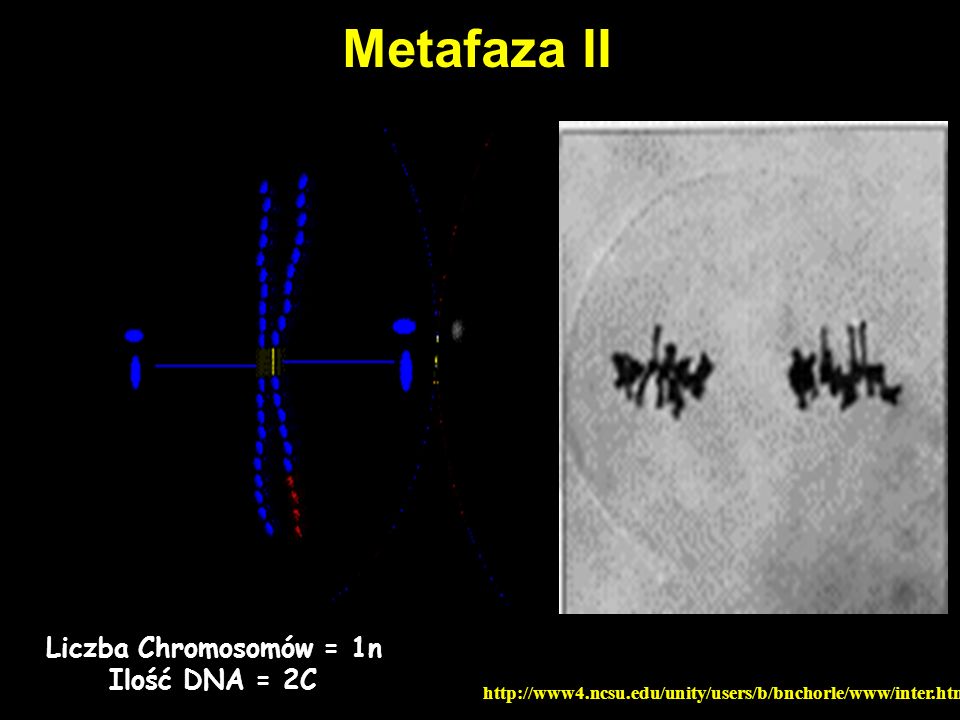
56
Anafaza II Liczba Chromosomów = 1n Ilość DNA = 1C

57
2-gi podział mejotyczny, to podział wyrównawczy
Telofaza II 1 2 3 4 2-gi podział mejotyczny, to podział wyrównawczy Liczba Chromosomów = 1n Ilość DNA = 1C

58
przyleganie mejotyczne
2n & 2C centromery widełki replikacyjne przyleganie mejotyczne Synaptotyczny kompleksotecomowe białka b tubule wrzeciona 2n & 4C 2x(1n & 2C) 4x(1n & 1C) Aleksander L. Sieroń
 4x(1n & 1C) Aleksander L. Sieroń.")
59
K O N I E C WYKŁADU 2 DZIĘKUJĘ

Podobne prezentacje





 ZALECANE PODRĘCZNIKI (do przygotowania do ćwiczeń i uzupełnienia.>")
>")












