Pobierz prezentację
1
Choroby genetyczne zwierząt Urszula Kosior-Korzecka
Przedmiot fakultatywny II rok weterynarii Urszula Kosior-Korzecka Zakład Patofizjologii Katedra Przedklinicznych Nauk Weterynaryjnych Wydział Medycyny Weterynaryjnej Uniwersytet Przyrodniczy w Lublinie

2
Program zajęć Ćwiczenie 1.
Ćwiczenie 1. Mechanizmy zaburzeń epigenetycznych, autosomalnych recesywnych i dominujących oraz sprzężonych z płcią. Mechanizmy dziedziczenia chorób autosomalnych i sprzężonych z płcią. Zależność fenotyp-genotyp w chorobach monogenowych na przykładzie talasemii i SCID-X. Sposoby postępowania ze zwierzętami chorymi i eliminacja nosicieli. Choroby genetyczne zwierząt, ważne z klinicznego i gospodarczego punktu widzenia Analiza mikromacierzy DNA, jako nowe narzędzie seryjnego badania ekspresji genów. Polimorfizm antygenów I i II klasy MHC a odporność i podatność na wybrane choroby infekcyjne i pasożytnicze. Białka histonowe w naprawie uszkodzonego DNA. SNP (single nucleotide polymorphism) - najnowsza metoda diagnozowania chorób genetycznych Ćwiczenie 2. Choroby genetyczne zwierząt powodowane przez mutacje chromosomalne strukturalne i liczbowe. - Morfologia i diploidalna liczba chromosomów u wybranych gatunków zwierząt gospodarskich i towarzyszących; typy mutacji chromosomalnych strukturalnych i liczbowych występujące u zwierząt; częstość występowania chorób warunkowanych przez mutacje chromosomowe strukturalne u zwierząt hodowlanych w Polsce; oznaczanie kariotypu i zasady opisu kariogramu; zastosowanie badań kariotypu u zwierząt gospodarskich. Etiologia i patogeneza wybranych chorób warunkowanych przez mutacje chromosomowe strukturalne u zwierząt: translokacja robertsonowska u krów, owiec, świń i lisów. Odwrócenie płci u psów i koni. Choroby uwarunkowane przez mutacje chromosomowe liczbowe u zwierząt: Monosomia chromosomu X u klaczy. Trisomie autosomalne u bydła i koni. Zaburzenia dojrzewania jądrowego oocytów u bydła i świń prowadzace do zaburzeń chromosomalnych liczbowych (powstawanie oocytów diploidalnych). Znaczenie mutacji chromosomalnych strukturalnych w patogenezie chorób nowotworowych u zwierząt. Uniparentalna disomia oraz imprinting genomowy w patogenezie aberracji chromosomowych u zwierząt. - Izolacja i zakładanie hodowli limfocytów, indukcja proliferacji w hodowli limfocytów – przygotowanie hodowli do analizy kariotypu – ćwiczenie praktyczne. - Oznaczanie kariotypu świni w dzielących się mitotycznie (PHA) limfocytach krwi –ćwiczenie praktyczne). - Analiza liczby chromosomów w oocytach świń hodowanych in vitro w celu eliminacji oocytów diploidalnych - ćwiczenie praktyczne.
 - najnowsza metoda diagnozowania chorób genetycznych. Ćwiczenie 2. Choroby genetyczne zwierząt powodowane przez mutacje chromosomalne strukturalne i liczbowe. - Morfologia i diploidalna liczba chromosomów u wybranych gatunków zwierząt gospodarskich i towarzyszących; typy mutacji chromosomalnych strukturalnych i liczbowych występujące u zwierząt; częstość występowania chorób warunkowanych przez mutacje chromosomowe strukturalne u zwierząt hodowlanych w Polsce; oznaczanie kariotypu i zasady opisu kariogramu; zastosowanie badań kariotypu u zwierząt gospodarskich. Etiologia i patogeneza wybranych chorób warunkowanych przez mutacje chromosomowe strukturalne u zwierząt: translokacja robertsonowska u krów, owiec, świń i lisów. Odwrócenie płci u psów i koni. Choroby uwarunkowane przez mutacje chromosomowe liczbowe u zwierząt: Monosomia chromosomu X u klaczy. Trisomie autosomalne u bydła i koni. Zaburzenia dojrzewania jądrowego oocytów u bydła i świń prowadzace do zaburzeń chromosomalnych liczbowych (powstawanie oocytów diploidalnych). Znaczenie mutacji chromosomalnych strukturalnych w patogenezie chorób nowotworowych u zwierząt. Uniparentalna disomia oraz imprinting genomowy w patogenezie aberracji chromosomowych u zwierząt. - Izolacja i zakładanie hodowli limfocytów, indukcja proliferacji w hodowli limfocytów – przygotowanie hodowli do analizy kariotypu – ćwiczenie praktyczne. - Oznaczanie kariotypu świni w dzielących się mitotycznie (PHA) limfocytach krwi –ćwiczenie praktyczne). - Analiza liczby chromosomów w oocytach świń hodowanych in vitro w celu eliminacji oocytów diploidalnych - ćwiczenie praktyczne.")
3
Ćwiczenie 3. Etiologia i patogeneza autosomalnych chorób genetycznych u zwierząt warunkowanych przez mutacje genowe. Typy mutacji genowych. Etiologia i patogeneza wybranych chorób monogenowych autosomalnych recesywnych u zwierząt: BLAD; CLAD; DUMPS; PSS; niedokrwistość hemolityczna u psów wywołana niedoborem kinazy pirogronianowej; choroba Andersena u kotów; mukopolisacharydoza u psów i kotów. Etiologia i patogeneza wybranych chorób monogenowych autosomalnych dominujących u zwierząt: hipercholesterolemia u świń i królików; porfiria u świń. Choroby powodowane przez mutacje autosomalne z niepełną dominacją: sferocytoza u bydła; choroba von Willebranda u dobermanów (typ IB). Techniki biologii molekularnej umożliwiające diagnozowanie i/lub wykrywanie nosicielstwa choroby monogenowej - PCR, RFLP, Real-Time PCR. Wykorzystanie technik PCR i RFLP do wykrywania nosicielstwa na przykładzie BLAD, DUMPS i PSS. Izolacja DNA z limfocytów krów, amplifikacja fragmentu genu podjednostki CD18 beta-integryny metodą PCR i elektroforetyczny rozdział produktów amplifikacji w żelu agarozowym – ćwiczenie praktyczne. Ćwiczenie 4. Choroby monogenowe sprzężone z płcią. Chromosomy płci u ssaków i ptaków; dziedziczenie i determinacja płci u ssaków i ptaków; inaktywacja chromosomu X; determinacja płci przy udziale genów autosomalnych oraz zlokalizowanych w chromosomach płci; cechy sprzężone z płcią. Etiologia i patogeneza chorób genetycznych monogenowych sprzężonych z płcią u zwierząt: zespół Alporta u psów; pierwotna hiperoksaluria u kotów; ograniczenie owulacji u kur; SCID-X u psów; zespół Menkesa u psów i owiec; dystrofie mięśniowe u psów i kotów; hemofilia A u koni, psów, kotów, świń i owiec; hemofilia B u psów i kotów. Wykorzystanie analizy rodowodów w eliminacji chorób sprzężonych z płcią w stadzie - zadania genetyczne. Zaburzenia procesu formowania się płci. Wybarwianie chromosomów płci w komórkach nabłonkowych lub komórkach krwi świń - ćwiczenie praktyczne
. Techniki biologii molekularnej umożliwiające diagnozowanie i/lub wykrywanie nosicielstwa choroby monogenowej - PCR, RFLP, Real-Time PCR. Wykorzystanie technik PCR i RFLP do wykrywania nosicielstwa na przykładzie BLAD, DUMPS i PSS. Izolacja DNA z limfocytów krów, amplifikacja fragmentu genu podjednostki CD18 beta-integryny metodą PCR i elektroforetyczny rozdział produktów amplifikacji w żelu agarozowym – ćwiczenie praktyczne. Ćwiczenie 4. Choroby monogenowe sprzężone z płcią. Chromosomy płci u ssaków i ptaków; dziedziczenie i determinacja płci u ssaków i ptaków; inaktywacja chromosomu X; determinacja płci przy udziale genów autosomalnych oraz zlokalizowanych w chromosomach płci; cechy sprzężone z płcią. Etiologia i patogeneza chorób genetycznych monogenowych sprzężonych z płcią u zwierząt: zespół Alporta u psów; pierwotna hiperoksaluria u kotów; ograniczenie owulacji u kur; SCID-X u psów; zespół Menkesa u psów i owiec; dystrofie mięśniowe u psów i kotów; hemofilia A u koni, psów, kotów, świń i owiec; hemofilia B u psów i kotów. Wykorzystanie analizy rodowodów w eliminacji chorób sprzężonych z płcią w stadzie - zadania genetyczne. Zaburzenia procesu formowania się płci. Wybarwianie chromosomów płci w komórkach nabłonkowych lub komórkach krwi świń - ćwiczenie praktyczne.")
4
Ćwiczenie 5. Etiologia i patogeneza wybranych chorób genetycznych u bydła i koni genetyczne podstawy rozwoju BSE wieloczynnikowe i jednoczynnikowe choroby genetyczne koni genetyczna podatność na nowotwory u koni o siwym umaszczeniu (czerniaki) markery chorób nowotworowych u koni Etiologia i patogeneza chorób genetycznych u kotów i psów. struktura genomu psów a podatność na wybrane choroby rasowa podatność genetyczna na rozwój cukrzycy I i II typu sercowo-naczyniowe i hematologiczne choroby genetyczne neurologiczne choroby genetyczne choroby przewodu pokarmowego o podłożu genetycznym testy DNA do rozpoznawania chorób genetycznych u psów oznaczanie ryzyka chorób genetycznych psów w oparciu o analizę rodowodową na przykładzie dysplazji stawu biodrowego Ćwiczenie 6. Etiologia i patogeneza chorób genetycznych świń, owiec i kóz. Uwarunkowania genetyczne determinujące podatność na rozwój chorób nowotworowych, plenność i wydajność mięsną świń, owiec i kóz. - manipulacje w genie IGF2 jako sposób poprawy wydajności mięsnej świń genetyczna oporność i podatność owiec na chłoniaki genetyczne uwarunkowania wielokrotnych owulacji u owiec hipercholesterolemia u świń rodzinne wole u owiec i kóz Dziedziczne niedobory immunologiczne u zwierząt. Genetyczna kontrola odporności na choroby. genetyczne podstawy odporności wrodzonej przykłady dziedzicznych niedoborów immunologicznych Transformacja blastyczna limfocytów wyizolowanych z krwi psa zdrowego i psa ze SCID-X in vitro – ćwiczenie praktyczne.
 markery chorób nowotworowych u koni. Etiologia i patogeneza chorób genetycznych u kotów i psów. struktura genomu psów a podatność na wybrane choroby. rasowa podatność genetyczna na rozwój cukrzycy I i II typu. sercowo-naczyniowe i hematologiczne choroby genetyczne. neurologiczne choroby genetyczne. choroby przewodu pokarmowego o podłożu genetycznym. testy DNA do rozpoznawania chorób genetycznych u psów. oznaczanie ryzyka chorób genetycznych psów w oparciu o analizę rodowodową na przykładzie dysplazji stawu biodrowego. Ćwiczenie 6. Etiologia i patogeneza chorób genetycznych świń, owiec i kóz. Uwarunkowania genetyczne determinujące podatność na rozwój chorób nowotworowych, plenność i wydajność mięsną świń, owiec i kóz. - manipulacje w genie IGF2 jako sposób poprawy wydajności mięsnej świń. genetyczna oporność i podatność owiec na chłoniaki. genetyczne uwarunkowania wielokrotnych owulacji u owiec. hipercholesterolemia u świń. rodzinne wole u owiec i kóz. Dziedziczne niedobory immunologiczne u zwierząt. Genetyczna kontrola odporności na choroby. genetyczne podstawy odporności wrodzonej. - przykłady dziedzicznych niedoborów immunologicznych. Transformacja blastyczna limfocytów wyizolowanych z krwi psa zdrowego i psa ze SCID-X in vitro – ćwiczenie praktyczne.")
5
Ćwiczenie 7. Genetyczne podstawy nowotworzenia aneuploidalność w komórkach nowotworowych psów znaczenie mutacji w protoonkogenach i genach supresorowych w rozwoju wybranych nowotworów u zwierząt gospodarskich i towarzyszących znaczenie translokacji wzajemnej w patomechanizmach rozwoju białaczek depresja inbredowa Terapia chorób uwarunkowanych genetycznie. Stosowane i aktualnie opracowywane terapie genowe; Zwierzęta transgeniczne jako modele chorób występujących u człowieka; Zwierzęta transgeniczne w produkcji biopreparatów. Ćwiczenie 8. Test zaliczeniowy

6
Choroba genetyczna to upośledzające sprawność życiową odchylenie od stanu prawidłowego, które przekazywane jest jako cecha dziedziczna z pokolenia na pokolenie, lub które powstaje de novo na skutek zmian i zaburzeń w mechanizmach przekazywania cech dziedzicznych (i może być również przekazywane potomstwu, jako cecha dziedziczna).
.")
7
Obecnie znamy około 3500 różnych chorób genetycznych.
Dzielimy je na 3 główne kategorie: · jednogenowe - będące wynikiem mutacji w jednym tylko genie. Choroby jednogenowe przekazywane są zgodnie z prawami Mendla i uwarunkowane treścią informacyjną jednego genu, tj. w obrębie pary alleli, występujących w określonym locus genowym. Wśród tych chorób wyróżnia się autosomalne recesywne, autosomalne dominujące i sprzężone z chromosomem płciowym żeńskim X (określane, jako sprzężone z płcią); · wielogenowe - będące wynikiem mutacji w wielu genach, warunkowane współdziałaniem wielu genów umiejscowionych w wielu loci. Choroby te nie są przekazywane zgodnie z prostym dziedziczeniem wg schematu Mendla. Niejednokrotnie objawy tych chorób występują na skutek interakcji z czynnikami środowiska i ujawniają się dopiero wówczas, gdy nasilenie działania tych czynników osiągnie pewną wartość progową; · chromosomowe - będące wynikiem mutacji na poziomie całych pakietów informacji genetycznej.
; · wielogenowe - będące wynikiem mutacji w wielu genach, warunkowane współdziałaniem wielu genów umiejscowionych w wielu loci. Choroby te nie są przekazywane zgodnie z prostym dziedziczeniem wg schematu Mendla. Niejednokrotnie objawy tych chorób występują na skutek interakcji z czynnikami środowiska i ujawniają się dopiero wówczas, gdy nasilenie działania tych czynników osiągnie pewną wartość progową; · chromosomowe - będące wynikiem mutacji na poziomie całych pakietów informacji genetycznej.")
8
Jednostkami chorobowymi łączonymi z mutacjami w mtDNA są:
- neuropatia nerwu wzrokowego; -encefalopatia mitochondrialna z kwasicą mleczanową i epizodami udaropodobnymi; - dziedziczna mateczna miopatia; - niektóre typy kardiomiopatii; -ataksja neurogenna; -cukrzyca insulinoniezależna; - cukrzyca insulinozależna; - barwnikowe zwyrodnienie siatkówki; - choroby neurodegeneracyjne wieku podeszłego.

10
Epigenetyczne dziedziczenie chorób

11
SCID-X Ciężki, sprzężony z płcią, złożony niedobór odporności u psów (SCID-X) Defekt genetyczny: mutacja w genie kodującym łańcuch gamma białkowego receptora interleukiny 2. IL-2 stanowi czynnik wzrostu dla limfocytów T i B, wzmaga aktywność komórek NK oraz przyspiesza dojrzewanie tymocytów. Delecja 4 pz, peptyd 21 aa (prawidłowy – 373 aa). Dziedziczenie: recesywne, sprzężone z chromosomem X. Rasy: Welsh Corgi, basset
 Defekt genetyczny: mutacja w genie kodującym łańcuch gamma białkowego receptora interleukiny 2. IL-2 stanowi czynnik wzrostu dla limfocytów T i B, wzmaga aktywność komórek NK oraz przyspiesza dojrzewanie tymocytów. Delecja 4 pz, peptyd 21 aa (prawidłowy – 373 aa). Dziedziczenie: recesywne, sprzężone z chromosomem X. Rasy: Welsh Corgi, basset")
12
SCID-X Cechy patologiczne:
mała dysplastyczna grasica (chorych psów osiąga 10% prawidłowej wielkości); zanik węzłów chłonnych; obniżona liczba limfocytów we krwi; brak lub znaczące obniżenie blastogennej odpowiedzi limfocytów na mitogeny i specyficzne antygeny - limfocyty T nie ulegają transformacji blastycznej pod wpływem PHA (fitohemaglutyniny), a limfocyty B pod wpływem miogenu ze szkarłatki.; hypogammaglobulinemia lub agammaglobulinemia. Poziom IgM w surowicy może być prawidłowy, ale poziom IgG jest dramatycznie niższy. Zdarza się, że objawy choroby pojawiają się u suk nosicielek, jeżeli zmutowany gen znajduje się na aktywnym chromosomie X (próby leczenia ludzką IL-2).
; zanik węzłów chłonnych; obniżona liczba limfocytów we krwi; brak lub znaczące obniżenie blastogennej odpowiedzi limfocytów na mitogeny i specyficzne antygeny - limfocyty T nie ulegają transformacji blastycznej pod wpływem PHA (fitohemaglutyniny), a limfocyty B pod wpływem miogenu ze szkarłatki.; hypogammaglobulinemia lub agammaglobulinemia. Poziom IgM w surowicy może być prawidłowy, ale poziom IgG jest dramatycznie niższy. Zdarza się, że objawy choroby pojawiają się u suk nosicielek, jeżeli zmutowany gen znajduje się na aktywnym chromosomie X (próby leczenia ludzką IL-2).")
13
Niedobory enzymatyczne szlaku glikolitycznego
Jest to grupa dziedzicznych enzymopatii. Ich przyczynę stanowi niedobór lub brak któregoś z enzymów szlaku glikoli tycznego w erytrocytach. Wspólnym objawem tych chorób jest NIEDOKRWISTOŚĆ HEMOLITYCZNA związana z drastycznie skrócona długością życia erytrocytów, retikulocytozą i żółtaczką.

14
Niedokrwistość hemolityczna u psów wywołana niedoborem kinazy pirogronianowej
Defekt genetyczny: mutacja w genie kodującym kinazę pirogronianową (PK). Dziedziczenie: autosomalne recesywne. Rasy: basenji, beagle, teriery.
. Dziedziczenie: autosomalne recesywne. Rasy: basenji, beagle, teriery.")
15
Niedobory enzymatyczne szlaku glikolitycznego
Niedokrwistość hemolityczna u psów wywołana niedoborem kinazy pirogronianowej. Defekt genetyczny: mutacja w genie kodującym kinazę pirogronianową (PK). Niedobór tego enzymu upośledza znacznie proces glikolizy w erytrocytach. Powoduje to zmniejszenie syntezy ATP, co w konsekwencji narusza integralność błony erytrocytów. (PK jest jedynym źródłem NADPH w erytrocytach koniecznego do redukcji glutationu. Zredukowany glutation jest ważnym antyoksydantem chroniącym przed RFT lipidy i białka błony komórkowej. Przy jego braku lub niedoborze, narasta stres oksydatywny i dochodzi do uszkodzeń błony i utraty jej integralności. Dziedziczenie: autosomalne recesywne.
. Niedobór tego enzymu upośledza znacznie proces glikolizy w erytrocytach. Powoduje to zmniejszenie syntezy ATP, co w konsekwencji narusza integralność błony erytrocytów. (PK jest jedynym źródłem NADPH w erytrocytach koniecznego do redukcji glutationu. Zredukowany glutation jest ważnym antyoksydantem chroniącym przed RFT lipidy i białka błony komórkowej. Przy jego braku lub niedoborze, narasta stres oksydatywny i dochodzi do uszkodzeń błony i utraty jej integralności. Dziedziczenie: autosomalne recesywne.")
16
Niedobory enzymatyczne szlaku glikolitycznego
Niedokrwistość hemolityczna u psów wywołana niedoborem kinazy pirogronianowej. Objawy: powiększenie obwodu brzucha, powiększenie śledziony, osłabienie , żółtaczka, zwiększenie stężenia pośredniej bilirubiny, skrócenie czasu życia erytrocytów, retykulocytoza, hemoglobinemia, niskie stężenie haptoglobiny w osoczu (tworzy ona kompleks z produktami rozpadu Hb, zwłaszcza z hemem, który usuwany jest przez watrobę, co stanowi ochronę dla nerek), wtórna hemochromatoza, zwłóknienie szpiku i stwardnienie kości. Krwinki z niedoborem PK są bardziej podatne na osmolizę (rozpad w warunkach obniżonego ciśnienia osmotycznego). Nosiciele charakteryzują się częściowym niedoborem PK. Rasy: basenji, beagle, teriery. Od 1995 r. istnieje test genetyczny w kierunku wykrywania psów z niedoborem PK opracowany przez Whitney i wsp.
, wtórna hemochromatoza, zwłóknienie szpiku i stwardnienie kości. Krwinki z niedoborem PK są bardziej podatne na osmolizę (rozpad w warunkach obniżonego ciśnienia osmotycznego). Nosiciele charakteryzują się częściowym niedoborem PK. Rasy: basenji, beagle, teriery. Od 1995 r. istnieje test genetyczny w kierunku wykrywania psów z niedoborem PK opracowany przez Whitney i wsp.")
17
Hiperlipoproteinemia I u psów. Niedobór lipazy lipoproteinowej
Hiperlipoproteinemia I u psów. Niedobór lipazy lipoproteinowej. Hipertriacyloglicerolemia. Hiperchylomikronemia. Defekt genetyczny: mutacja w genie kodującym lipazę lipoproteinową. Dziedziczenie: autosomalne recesywne. Za transport wszystkich lipidów (zawartych w pokarmie) do układu krążenia odpowiedzialne są chylomikrony. Lipaza lipoproteinowa katalizuje rozpad chylomikronów oraz VLDL do tzw. resztkowych lipoprotein. Istnieje dodatnia korelacja pomiędzy aktywnością LPL a zdolnością tkanek do wbudowywania kwasów tłuszczowych z triacylogliceroli Lp.
 do układu krążenia odpowiedzialne są chylomikrony. Lipaza lipoproteinowa katalizuje rozpad chylomikronów oraz VLDL do tzw. resztkowych lipoprotein. Istnieje dodatnia korelacja pomiędzy aktywnością LPL a zdolnością tkanek do wbudowywania kwasów tłuszczowych z triacylogliceroli Lp.")
18
Hiperlipoproteinemia I u psów. Niedobór lipazy lipoproteinowej
Hiperlipoproteinemia I u psów. Niedobór lipazy lipoproteinowej. Hipertriacyloglicerolemia. Hiperchylomikronemia.

19
Hiperlipoproteinemia I u psów. Niedobór lipazy lipoproteinowej
Hiperlipoproteinemia I u psów. Niedobór lipazy lipoproteinowej. Hipertriacyloglicerolemia. Hiperchylomikronemia. Dziedziczny niedobór LPL charakteryzuje się bardzo powolnym klirensem chylomikronów z krążenia, co prowadzi do odbiegających od normy dużych stężeń chylomikronów w osoczu. Odnotowuje się także: - wysokie stężenie endogennych triacylogliceroli (obecnych w VLDL); - zmniejszone stężenie frakcji LDL i HDL. Ten stan może być dodatkowo indukowany dietą wysokotłuszczową. Koryguje się go dietą o małej zawartości tłuszczu i zwiększeniem w karmie ilości złożonych węglowodanów. U osobników homozygotycznych recesywnych stwierdza się całkowity brak enzymu, natomiast u osobników heterozygotycznych zachodzi prawidłowa lipoliza chylomikronów. Stwierdza się odkładanie chylomikronów w makrofagach skóry, komórkach Browicza-Kupfera i rogówce. Rasy: Beagle
; - zmniejszone stężenie frakcji LDL i HDL. Ten stan może być dodatkowo indukowany dietą wysokotłuszczową. Koryguje się go dietą o małej zawartości tłuszczu i zwiększeniem w karmie ilości złożonych węglowodanów. U osobników homozygotycznych recesywnych stwierdza się całkowity brak enzymu, natomiast u osobników heterozygotycznych zachodzi prawidłowa lipoliza chylomikronów. Stwierdza się odkładanie chylomikronów w makrofagach skóry, komórkach Browicza-Kupfera i rogówce. Rasy: Beagle")
20
ZESPÓŁ ALPORTA Defekt genetyczny: Mutacja w 35 eksonie genu (COL4A5) kodującego łańcuch alfa 5 kolagenu typu IV. Skutkiem mutacji jest nieprawidłowa budowa domeny Nc1 (skrócenie sekwencji aminokwasowej) białka kolagenu typu IV. Dziedziczenie: recesywne, sprzężone z płcią. Z powodu kojarzenia zwierząt w pokrewieństwie i późne pojawienie się objawów choroba ta może wystąpić także u suk, homozygotycznych pod względem zmutowanego genu. Suki jednak nie giną, chociaż ulegają znacznemu wyniszczeniu. Przyczyną choroby jest nieprawidłowa budowa białka kolagenu IV. Kolagen typu IV znajduje się w kłębkach nerkowych oraz błonach podstawowych wszystkich komórek nabłonkowych i śródbłonkowych.
 kodującego łańcuch alfa 5 kolagenu typu IV. Skutkiem mutacji jest nieprawidłowa budowa domeny Nc1 (skrócenie sekwencji aminokwasowej) białka kolagenu typu IV. Dziedziczenie: recesywne, sprzężone z płcią. Z powodu kojarzenia zwierząt w pokrewieństwie i późne pojawienie się objawów choroba ta może wystąpić także u suk, homozygotycznych pod względem zmutowanego genu. Suki jednak nie giną, chociaż ulegają znacznemu wyniszczeniu. Przyczyną choroby jest nieprawidłowa budowa białka kolagenu IV. Kolagen typu IV znajduje się w kłębkach nerkowych oraz błonach podstawowych wszystkich komórek nabłonkowych i śródbłonkowych.")
21
ZESPÓŁ ALPORTA Objawy pojawiają się dopiero około 4 roku życia.
Występuje brak apetytu, chudnięcie i wymioty. Następnie stwierdza się: - niezagęszczanie moczu; - białkomocz; - krwiomocz; - zapalenie kłębków nerkowych; - niewydolność nerek; - postępujące wyniszczenie; - osłabienie słuchu.

22
Ultrastructural appearance of glomerular basement membrane from affected Dalmatians with autosomal dominant Alport syndrome: (A) normal membrane in 35‐day‐old embryo (×5700); (B) lamellated membrane with fusion of overlying foot processes in adult dog (×1850); (C) irregular thickened lamellated membrane with subepithelial frilling and fusion of foot processes in adult dog (×1850); and (D) membrane showing irregular basket weave appearance in adult dog (×5700). Ultrastructural appearance of glomerular basement membrane from affected Dalmatians with autosomal dominant Alport syndrome: (A) normal membrane in 35‐day‐old embryo (×5700); (B) lamellated membrane with fusion of overlying foot processes in adult dog (×1850); (C) irregular thickened lamellated membrane with subepithelial frilling and fusion of foot processes in adult dog (×1850); and (D) membrane showing irregular basket weave appearance in adult dog (×5700). Hood J C et al. Nephrol. Dial. Transplant. 2002;17: © European Renal Association–European Dialysis and Transplant Association
 normal membrane in 35‐day‐old embryo (×5700); (B) lamellated membrane with fusion of overlying foot processes in adult dog (×1850); (C) irregular thickened lamellated membrane with subepithelial frilling and fusion of foot processes in adult dog (×1850); and (D) membrane showing irregular basket weave appearance in adult dog (×5700). Hood J C et al. Nephrol. Dial. Transplant. 2002;17: © European Renal Association–European Dialysis and Transplant Association.")
23
ZESPÓŁ ALPORTA W badaniach mikroskopowych, po przeprowadzeniu biopsji nerek, stwierdza się: - atrofię kłębków nerkowych; - torbielowate rozszerzenie torebkowych przestrzeni Bowmana. Rokowanie jest złe, ale zgon można opóźnić przez zmianę diety na niskobiałkową. Rasy: bullterier i samojed.

24
ZESPÓŁ ALPORTA

25
Hemofilia B u psów i kotów (choroba Christmas).
Defekt genetyczny: tranzycja (mutacja zmiany sensu) A-G w nukleotydzie w genie kodującym IX czynnik krzepnięcia krwi. Prowadzi to do zamiany kwasu glutaminowego na glicynę w kodonie 379 w białku czynnika IX. Ta pojedyncza substytucja aminokwasowa znacząco zmienia strukturę trzeciorzędową cząsteczki czynnika IX, co prowadzi do utraty jego aktywności. Druga możliwość – delecje w genie kodującym czynnik IX. Dziedziczenie: recesywne, sprzężone z płcią.
 A-G w nukleotydzie 1477 w genie kodującym IX czynnik krzepnięcia krwi. Prowadzi to do zamiany kwasu glutaminowego na glicynę w kodonie 379 w białku czynnika IX. Ta pojedyncza substytucja aminokwasowa znacząco zmienia strukturę trzeciorzędową cząsteczki czynnika IX, co prowadzi do utraty jego aktywności. Druga możliwość – delecje w genie kodującym czynnik IX. Dziedziczenie: recesywne, sprzężone z płcią.")
26
Hemofilia B występuje rzadziej, niż hemofilia A.
Objawy są podobne. Hemofilię B opisano u 20 ras psów oraz u kotów. Częściej, niż w przypadku hemofilii A spotyka się przypadki choroby u suk. Jednakże w przeciwieństwie do hemofilii A, w której występują lekkie, średnie i ciężkie niedobory czynnika VIII, większość opisanych u zwierząt przypadków hemofilii B charakteryzuje mniejszy niż 5% poziom czynnika IX. Badania wykazały występowanie 2 grup pacjentów: - z nieoznaczalnym poziomem czynnika IX; z normalną ilością zmutowanego (nieaktywnego) czynnika IX. Ciężkość krwawienia zależy od typu mutacji (tranzycja, czy delecja), ale jest zwykle proporcjonalna do stopnia niedoboru czynnika IX.
 czynnika IX. Ciężkość krwawienia zależy od typu mutacji (tranzycja, czy delecja), ale jest zwykle proporcjonalna do stopnia niedoboru czynnika IX.")
27
Nasilenie objawów klinicznych hemofilii B w znacznym stopniu uzależnione jest od wielkości psa. (!!!!!!!!!!!!!!!!!!!!!!!!!!!) U małych ras (teriery, cocker spaniel, owczarek szetlandzki, buldog francuski) jest lekkie lub średnie. Ciężka postać występuje u dużych ras (bernardyn, Alaskan malamut, owczarek staro angielski, owczarek niemiecki). Hemofilia B objawia się głównie: - występowaniem krwiaków; - wylewami krwawymi do jam stawowych; - krwotokami z układu pokarmowego lub do jam ciała. W przypadku łagodnej formy psy osiągają dojrzałość płciową i mogą przekazać chorobę potomstwu. Diagnostyka podobna, z tą różnicą, że stwierdza się obniżony poziom czynnika IX.
 jest lekkie lub średnie. Ciężka postać występuje u dużych ras (bernardyn, Alaskan malamut, owczarek staro angielski, owczarek niemiecki). Hemofilia B objawia się głównie: - występowaniem krwiaków; - wylewami krwawymi do jam stawowych; - krwotokami z układu pokarmowego lub do jam ciała. W przypadku łagodnej formy psy osiągają dojrzałość płciową i mogą przekazać chorobę potomstwu. Diagnostyka podobna, z tą różnicą, że stwierdza się obniżony poziom czynnika IX.")
28
Niezgodność grup krwi i konflikt serologiczny u kotów
Przez lata hodowcy kotów borykali się z problemem niewyjaśnionej śmiertelności wśród kocich noworodków, która dotykała jedne rasy częściej od innych. Dopiero badania w latach 80-tych ubiegłego wieku nad grupami krwi u kotów wykazały, że jednym z powodów syndromu słabnących kociąt (Fading Kitten Syndrome) jest izoerytroliza noworodków, wywoływana przez niezgodność grupy krwi matki i jej potomstwa. Blue-eyed cats with white fur have a higher incidence of genetic deafness.
 jest izoerytroliza noworodków, wywoływana przez niezgodność grupy krwi matki i jej potomstwa. Blue-eyed cats with white fur have a higher incidence of genetic deafness.")
29
U kotów występują trzy grupy krwi: A (najbardziej powszechna); B (częsta u niektórych ras); AB (niezależnie od rasy występująca bardzo rzadko). Grupa krwi jest determinowana genetycznie inaczej, niż u człowieka. U kotów gen odpowiedzialny za grupę krwi A jest dominujący w stosunku do genu na grupę B. Oznacza to, że tylko kot posiadający dwa allele warunkujące grupę B będzie miał grupę krwi B. Kot z grupą krwi A może być zarówno homozygotą (posiadać dwa allele A), jak i heterozygotą (posiadać jeden allel A i jeden allel B). Trzeci rodzaj grupy krwi występującej u kotów, grupa AB, jest jeszcze nie w pełni poznany. Czynnikiem determinującym tę grupę krwi jest osobny allel , recesywny w stosunku do A, ale dominujący w stosunku do B.
, jak i heterozygotą (posiadać jeden allel A i jeden allel B). Trzeci rodzaj grupy krwi występującej u kotów, grupa AB, jest jeszcze nie w pełni poznany. Czynnikiem determinującym tę grupę krwi jest osobny allel , recesywny w stosunku do A, ale dominujący w stosunku do B.")
30
Grupa krwi A występuje najczęściej w populacji kotów domowych zarówno krótko-, jak i długowłosych, przy czym u tych osobników bardzo rzadko stwierdzano w surowicy duże stężenie naturalnych przeciwciał. Odwrotną sytuację stwierdzono u dużo mniejszej liczby kotów z grupą krwi B, u których bardzo często w surowicy krwi występuje duże stężenie naturalnych przeciwciał anty-A. Wśród kotów rasowych stwierdza się znaczne różnice w częstości występowania określonych grup krwi z powodu stosowania długotrwałej selekcji skierowanej na różne cechy. U kotów w razie przetoczenia krwi niezgodnej grupowo okres życia erytrocytów znacznie się skraca i trwa od kilku godzin do kilku dni, podczas gdy normalna długość życia krwinek czerwonych u kotów wynosi 4-5 tygodni. Transfuzje takie mogą wywołać ostrą reakcję organizmu, szczególnie, gdy erytrocyty z grupą A zostały (nawet pierwszy raz) zmieszane z krwinkami grupy B.
 zmieszane z krwinkami grupy B.")
31
Identyfikacja grup krwi u kotów i psów umożliwia rozpoznanie i zapobieganie zjawisku ERYTROLIZY NOWORODKÓW. Do izoelektrolizy, czyli niedokrwistości hemolitycznej dochodzi, gdy nowonarodzone kocieta posiadają inną grupę krwi (A lub AB,) niż matka (B). Ssac siarę otrzymują jednocześnie przeciwciała skierowane przeciwko własnym czerwonym krwinkom, które są niszczone, gdy tylko przeciwciała dostaną się do krwiobiegu kociąt. Choroba ta jest największym problemem w hodowli kotów – tzw. syndrom słabnących kociąt, u których często występuje grupa B. Do erytrolizy dochodzi wtedy, gdy kocięta z grupą krwi A (pochodzące od ojca z grupą A) ssą siarę od matki z grupą B. W siarze kotek z grupą krwi B znajdują się naturalne przeciwciałą anty-A, które po wchłonięciu się z przewodu pokarmowego kociąt wiążą się z ich erytrocytami, wywołując: - żółtaczkę, - bilirubinurię, - niedokrwistość , - śmierć w ciągu kilku dni.
 niż matka (B). Ssac siarę otrzymują jednocześnie przeciwciała skierowane przeciwko własnym czerwonym krwinkom, które są niszczone, gdy tylko przeciwciała dostaną się do krwiobiegu kociąt. Choroba ta jest największym problemem w hodowli kotów – tzw. syndrom słabnących kociąt, u których często występuje grupa B. Do erytrolizy dochodzi wtedy, gdy kocięta z grupą krwi A (pochodzące od ojca z grupą A) ssą siarę od matki z grupą B. W siarze kotek z grupą krwi B znajdują się naturalne przeciwciałą anty-A, które po wchłonięciu się z przewodu pokarmowego kociąt wiążą się z ich erytrocytami, wywołując: - żółtaczkę, - bilirubinurię, - niedokrwistość , - śmierć w ciągu kilku dni.")
32
Krzyżówka przedstawiająca dziedziczenie grup krwi u kotów
Kotka Grupa krwi A Grupa krwi B Genotyp: AA Ab bb Kocur Grupa krwi A AA - grupa A AA - grupa A Ab - grupa A Ab - grupa A Konflikt Ab AA - grupa A Ab - grupa A bb - grupa B Ab - grupa A Konflikt bb - grupa B bb Ab - grupa A Ab - grupa A bb - grupa B bb - grupa B

33
A litter of kittens with neonatal isoerythrolysis
A litter of kittens with neonatal isoerythrolysis. Note the jaundiced (yellow) noses and bloody tip of the tail of the white kitten.
 noses and bloody tip of the tail of the white kitten.")
34
Okresowe porażenie u koni „quarter” związany z nadmiarem potasu
Okresowe porażenie u koni „quarter” związany z nadmiarem potasu. HYPP – hyperkalemic periodic paralysis. Defekt genetyczny: mutacja typu transwersja (C-G) w regionie S3 podjednostki genu, prowadząca do zmiany sekwencji aminokwasów (Phe-Leu) – zmaina sensu; w białkowym kanale sodowym komórek dojrzałych mięśni szkieletowych. Istnieje test genetyczny umożliwiający wykrywanie heterozygot. Dziedziczenie: autosomalne dominujące. Chorują głównie heterozygoty, gdyż homozygoty dominujące we wczesnym wieku źrebięcym giną z powodu trudności z oddychaniem i porażeniem dużych partii mięśni. Choroba ta występuje najczęściej u koni amerykańskich biegających na ćwierć mili. Objawy to: okresowe pęczkowe drżenie i skurcze mięśni, osłabienie, skłonność do przyjmowania pozycji leżącej oraz nadmiar potasu we krwi i czasem niewydolność oddechowa, związana z porażeniem mięśni krtani i gardła. Objawy mogą wystąpić lub nasilić się po podaniu paszy bogatej w potas (lucerna). Potencjalne następstwa ataków to otarcia skóry, bezwiedne upadki i związane z tym kontuzje. Okresowy paraliż obejmuje podczas ataku duże partie mięśni konia, a ponieważ większość opisanych przypadków z lekkimi objawami to osobniki heterozygotyczne, sugeruje to, że osobniki homozygotyczne pod względem tej dominującej anomalii mogą mieć znacznie bardziej nasilone objawy. Ponadto u osobników obarczonych tą anomalią podczas stosowania anestezji halotanowej mogą wystąpić objawy hipertermii złośliwej. W leczeniu stosuje się fenytoinę, która działa jako środek antykonwulsyjny i jest skuteczna przez długi okres po jej zastosowaniu.
 w regionie S3 podjednostki genu, prowadząca do zmiany sekwencji aminokwasów (Phe-Leu) – zmaina sensu; w białkowym kanale sodowym komórek dojrzałych mięśni szkieletowych. Istnieje test genetyczny umożliwiający wykrywanie heterozygot. Dziedziczenie: autosomalne dominujące. Chorują głównie heterozygoty, gdyż homozygoty dominujące we wczesnym wieku źrebięcym giną z powodu trudności z oddychaniem i porażeniem dużych partii mięśni. Choroba ta występuje najczęściej u koni amerykańskich biegających na ćwierć mili. Objawy to: okresowe pęczkowe drżenie i skurcze mięśni, osłabienie, skłonność do przyjmowania pozycji leżącej oraz nadmiar potasu we krwi i czasem niewydolność oddechowa, związana z porażeniem mięśni krtani i gardła. Objawy mogą wystąpić lub nasilić się po podaniu paszy bogatej w potas (lucerna). Potencjalne następstwa ataków to otarcia skóry, bezwiedne upadki i związane z tym kontuzje. Okresowy paraliż obejmuje podczas ataku duże partie mięśni konia, a ponieważ większość opisanych przypadków z lekkimi objawami to osobniki heterozygotyczne, sugeruje to, że osobniki homozygotyczne pod względem tej dominującej anomalii mogą mieć znacznie bardziej nasilone objawy. Ponadto u osobników obarczonych tą anomalią podczas stosowania anestezji halotanowej mogą wystąpić objawy hipertermii złośliwej. W leczeniu stosuje się fenytoinę, która działa jako środek antykonwulsyjny i jest skuteczna przez długi okres po jej zastosowaniu.")
35
Choroba Krabbego u psów
Defekt genetyczny: mutacja w genie kodującym beta-galaktozydazę, znosząca lub zmniejszająca aktywność tego enzymu. Dziedziczenie: autosomalne recesywne. Jest to sfingolipidoza związana ze spichrzaniem w komórkach układu nerwowego galaktozyloceramidu. Galaktozyloceramid - złożony lipid, który nie ulega degradacji ze względu na brak aktywności enzymu lizosomalnego – beta-galaktozydazy, odpowiedzialnego za rozkład do beta-galaktozy oraz ceramidu. Brak degradacji galaktozyloceramidu prowadzi z jednej strony do gromadzenia w komórkach n ie rozłożonego galaktozyloceramidu, a z drugiej strony do braku ceramidu jako substratu do syntezy sfingozyny i sfingomieliny. W efekcie u osobników chorych występuje prawie całkowity brak mieliny.

36
Choroba Krabbego u psów
Objawy: początek choroby w młodym wieku, choroba poczatkowo ma charakter zaburzeń mentalnych, prowadzi do paraliżu i śmierci. Pierwsze objawy kliniczne leukodystrofii globoidalnej pojawiają się u dotkniętych nią psów w wieku 1-3 miesięcy. Należą do nich ataksja oraz porażenie tylnych kończyn. W dalszym przebiegu choroby następuje atrofia mięśni oraz degeneracja neurologiczna. Ze względu na brak możliwości leczenia chore zwierzęta są usypiane w wieku 10 miesięcy.

37
Choroba Krabbego u psów
W Wielkiej Brytanii u dorosłych białych terrierów West Highland przed dopuszczeniem do reprodukcji wykonywane są testy DNA w celu wykrywania i eliminacji choroby Krabbego w hodowlach.

38
Mukopolisacharydoza typu I u psów (i kotów)
Mukopolisacharydoza typu I u psów (i kotów). Niedobór alfa-L-iduronidazy. Zespół Hurler/Scheiego. Defekt genetyczny: mutacja w genie kodującym alfa-L-iduronidazę – enzym lizosomalny degradujący dwa z 7 metabolitów GAG. Dziedziczenie: autosomalne recesywne. Przyczyną choroby jest niedobór enzymu lizosomalnego alfa-L-iduronidazy, odpowiedzialnego za degradację GAG. Na skutek tego u chorych zwierząt, głównie w komórkach wątrobowych, dochodzi do spichrzania dwóch z t metabolitów GAG. Są to: siarczan dermatanu (DS.) oraz siarczan heparanu (HS). Oba te metabolity są obecne w moczu chorych z mukopoisacharydozą typu I, co jest wykorzystywane w diagnostyce.
. Niedobór alfa-L-iduronidazy. Zespół Hurler/Scheiego. Defekt genetyczny: mutacja w genie kodującym alfa-L-iduronidazę – enzym lizosomalny degradujący dwa z 7 metabolitów GAG. Dziedziczenie: autosomalne recesywne. Przyczyną choroby jest niedobór enzymu lizosomalnego alfa-L-iduronidazy, odpowiedzialnego za degradację GAG. Na skutek tego u chorych zwierząt, głównie w komórkach wątrobowych, dochodzi do spichrzania dwóch z t metabolitów GAG. Są to: siarczan dermatanu (DS.) oraz siarczan heparanu (HS). Oba te metabolity są obecne w moczu chorych z mukopoisacharydozą typu I, co jest wykorzystywane w diagnostyce.")
39
Dziedziczna głuchota u psów dalmatyńskich
Rodzaj defektu genetycznego nie jest znany. Choroba dziedziczona jest autosomalnie recesywnie. Objawy: - wrodzona głuchota; - anomalie barwy oczu (oczy niebieskie); - występowanie od urodzenia ciemnych łat (a nie charakterystycznych dla dalmatyńczyków plamek).
; - występowanie od urodzenia ciemnych łat (a nie charakterystycznych dla dalmatyńczyków plamek).")
40
Dziedziczna głuchota u psów dalmatyńskich
Częstość występowania dalmatyńczyków z dziedziczną głuchotą w Norwegii wynosi 3,6%, natomiast w USA jest znacznie wyższa (w Polsce – brak danych). Fakt ten tłumaczy się prowadzoną przez hodowców norweskich selekcją mającą na celu wykluczenie osobników z niebieskimi oczami i nietypowymi dla dalmatyńczyków łatami, ponieważ cechy te są niezgodne ze wzorcem rasy.
. Fakt ten tłumaczy się prowadzoną przez hodowców norweskich selekcją mającą na celu wykluczenie osobników z niebieskimi oczami i nietypowymi dla dalmatyńczyków łatami, ponieważ cechy te są niezgodne ze wzorcem rasy.")
41
Translokacja albo fuzja robertsonowska chromosomu 1/29 u krowy

42
Kariotyp (chromosomy w płytce metafazowej) pochodzące od:
klaczy (64,XX) ogiera (64,XY) klaczy z monosomią chromosomu X (63,X0) klaczy z trisomią chromosomu X (65,XXX)
 ogiera (64,XY) klaczy z monosomią chromosomu X (63,X0) klaczy z trisomią chromosomu X (65,XXX)")
43
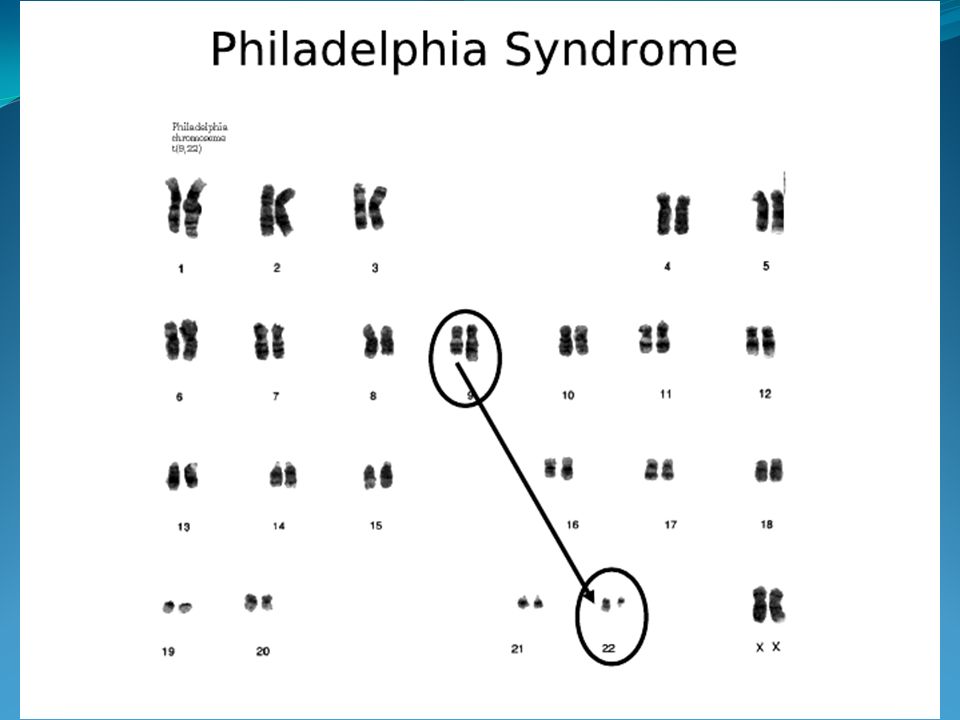
Genetyczne uwarunkowania przewlekłej białaczki szpikowej (translokacja wzajemna prowadząca do utworzenia chromosomu Philadelphia 1).
.")














 i WTÓRNE (NABYTE) NIEDOBORY ODPORNOŚCI>")






