Pobierz prezentację
This is a modal window.
1
Regulacja aktywności enzymów
Enzymologia-10 Regulacja aktywności enzymów

2
regulacji aktywności enzymów
Fizjologiczne sposoby regulacji aktywności enzymów Kontrola allosteryczna Modyfikacja kowalencyjna Działanie białek regulatorowych Aktywacja proteolityczna Działanie specyficznych inhibitorów Zmiany aktywności zależne od pH; kompartmentalizacja 7. Autoproteoliza

3
Kontrola allosteryczna
Małocząsteczkowe ligandy wiążąc się z enzymem oligomerycznym w określonym miejscu (centrum allosteryczne), powodują zmianę jego konformacji, a w efekcie – aktywności. Możliwe hamowanie lub zwiększanie aktywności. Regulacja płynna Hamowanie przez sprzężenie zwrotne
, powodują zmianę. jego konformacji, a w efekcie – aktywności. Możliwe hamowanie. lub zwiększanie aktywności. Regulacja płynna. Hamowanie przez sprzężenie zwrotne.")
4
Kontrola allosteryczna
Enzym karbamoilotransferaza asparaginianowa (ATCaza) katalizuje pierwszy etap biosyntezy pirymidyn, kondensację asparaginianu i karbamoilofosforanu CTP - końcowy produkt szlaku inicjowanego przez ATCazę – jest inhibitorem allosterycznym ATCazy
 katalizuje pierwszy etap. biosyntezy pirymidyn, kondensację asparaginianu i karbamoilofosforanu. CTP - końcowy produkt szlaku inicjowanego przez ATCazę – jest inhibitorem. allosterycznym ATCazy.")
5
Kontrola allosteryczna
ATCaza jest białkiem oligomerycznym. Jedna cząsteczka enzymu składa się z 12 podjednostek: dwóch trimerów podjednostek katalitycznych i trzech dimerów podjednostek regulatorowych.

6
Reakcja katalizowana przez karbamoilotransferazę asparaginianową Stan przejściowy Analog stanu przejściowego

7
Kontrola allosteryczna
ATCaza istnieje w dwóch stanach konformacyjnych. Stosunkowo zwarta forma T jest słabo aktywna katalitycznie, natomiast rozszerzona forma R jest w pełni aktywna. Związanie PALA, analogu stanu przejściowego reakcji katalizowanej przez ATCazę, stabilizuje formę R

8
Kontrola allosteryczna
W nieobecności substratów i/lub regulatorów allosterycznych ATCaza znajduje się w stanie równowagi pomiędzy formą T i R. W tych warunkach równowaga jest wyraźnie przesunięta w stronę formy T (około 200 ).
.")
9
Kontrola allosteryczna
Wiązanie CTP z podjednostka regulatorową ATCazy stabilizuje formę T

10
Kontrola allosteryczna
Enzymy allosteryczne wykazują kinetykę sigmoidalną Enzym allosteryczny można wyobrazić sobie jako mieszaninę dwóch enzymów wykazujących „klasyczną” kinetyke M-M, z których jeden wykazuje niskie powinowactwo do substratu (forma T), a drugi – wysokie (forma R). Ostateczna krzywa jest wynikiem „złożenia” dwóch krzywych cząstkowych. Przy niskich stężeniach substratu przeważa udział krzywej formy T; w miarę zwiększania [Asp] rośnie udział krzywej formy R.
, a drugi – wysokie (forma R). Ostateczna krzywa jest wynikiem. „złożenia dwóch krzywych cząstkowych. Przy niskich stężeniach substratu przeważa udział. krzywej formy T; w miarę zwiększania [Asp] rośnie udział krzywej formy R.")
11
Kontrola allosteryczna
Regulatory allosteryczne modulują stan równowagi pomiędzy formami R i T Inhibitor allosteryczny przesuwa równowagę w stronę formy T Aktywator allosteryczny przesuwa równowagę w stronę formy R Wiązanie regulatorów allosterycznych z ATCazą zmienia wartość L

12
MODELE ODDZIAŁYWAŃ ALLOSTERYCZNYCH
Model Hilla Wprowadzając: Y – stopień nasycenia; stosunek stężenia miejsc wiążących ligand zajętych przez cząsteczki liganda do ogólnego stężenia tych miejsc, czyli:

13
MODELE ODDZIAŁYWAŃ ALLOSTERYCZNYCH
n wyznaczone eksperymentalnie jest na ogół niższe niż rzeczywista liczba miejsc wiązania; n < 1 - ujemny efekt kooperatywny; n = 1 – brak efektu kooperatywnego; n > 1 – dodatni efekt kooperatywny

14
Model Monoda, Wymana i Changeaux
MODELE ODDZIAŁYWAŃ ALLOSTERYCZNYCH Model Monoda, Wymana i Changeaux (jednoprzejściowy) Założenia: podjednostki zajmują równoważne pozycje; co najmniej jedna oś symetrii konformacja podjednostki zależy od oddziaływań z innymi podjednostkami istnieją dwa możliwe stany konformacyjne oligomeru: R i T zmiana R T zachowuje symetrię oligomeru
 Założenia: podjednostki zajmują równoważne pozycje; co najmniej jedna oś symetrii. konformacja podjednostki zależy od oddziaływań z innymi podjednostkami. istnieją dwa możliwe stany konformacyjne oligomeru: R i T. zmiana R T zachowuje symetrię oligomeru.")
15
MODELE ODDZIAŁYWAŃ ALLOSTERYCZNYCH
Y – stopień nasycenia miejsc wiązania ligandami R – część cząsteczek enzymu w stanie R

16
MODELE ODDZIAŁYWAŃ ALLOSTERYCZNYCH
Model Koshlanda (sekwencyjny) Założenia: - wiązanie liganda z podjednostką zmienia jej konformację, co powoduje zmianę oddziaływań z inną podjednostką; - w nieobecności liganda występuje tylko jeden stan konformacyjny - zmiany konformacyjne są sekwencyjne - możliwy zarówno dodatni jak i ujemny efekt kooperatywny Wyniki badań wielu enzymów allosterycznych wskazują, że większość z nich zachowuje się zgodnie z modelem mieszanym, łączącym w sobie elementy modelu jednoprzejściowego i sekwencyjnego.
 Założenia: - wiązanie liganda z podjednostką zmienia jej konformację, co powoduje. zmianę oddziaływań z inną podjednostką; - w nieobecności liganda występuje tylko jeden stan konformacyjny. - zmiany konformacyjne są sekwencyjne. - możliwy zarówno dodatni jak i ujemny efekt kooperatywny. Wyniki badań wielu enzymów allosterycznych wskazują, że większość. z nich zachowuje się zgodnie z modelem mieszanym, łączącym w sobie. elementy modelu jednoprzejściowego i sekwencyjnego.")
17
Modyfikacja kowalencyjna

18
Modyfikacja kowalencyjna Fosforylacja i defosforylacja enzymów
Kinazy białkowe katalizują reakcje fosforylacji specyficznych reszt Ser, Thr lub Tyr. Fosfatazy białkowe katalizują reakcje hydrolitycznego usunięcia grup fosforanowych. Reszty Ser, Thr lub Tyr modyfikowane w wyniku działania kinaz białkowych wchodzą w skład specyficznych sekwencji rozpoznawanych przez kinazę. Kinazy i fosfatazy białkowe są aktywowane lub dezaktywowane przez małocząsteczkowe ligandy. Regulacja aktywności poprzez fosforylację/defosforylację ma często charakter 0/1, tzn. jedna z form regulowanego enzymu jest aktywna, a druga nie. Znane są jednak także przypadki, w których fosforylacja jedynie zwiększa albo zmniejsza aktywność enzymu lub w których zmienia podatność enzymu na działanie efektorów allosterycznych

19
Fosforylacja i defosforylacja enzymów
KLASYFIKACJA KINAZ BIAŁKOWYCH

20
Fosforylacja i defosforylacja enzymów
Zmiany konformacyjne fosforylazy glikogenowej zachodzące w wyniku fosforylacji

21
Fosforylacja i defosforylacja enzymów
Inaktywacja centrum aktywnego dehydrogenazy izocytrynianowej w wyniku fosforylacji Ser113

22
Fosforylacja i defosforylacja enzymów
Kinazy białkowe A są aktywowane przez cAMP Kinaza A rozpoznaje sekwencję -Arg-Arg-Gly-Ser-Ile- W podjednostce regulatorowej kinazy A występuje sekwencja przypominająca substrat -Arg-Arg-Gly-Ala-Ile- Mechanizm aktywacji kinazy białkowej A

23
Fosforylacja i defosforylacja enzymów
Wiązanie sekwencji przypominającej substrat z kinazą A Podjednostka katalityczna kinazy A związana z fragmentem przypominającym substrat podjednostki regulatorowej

24
Działanie białek regulatorowych
Aktywność niektórych enzymów zmienia się w wyniku oddziaływania z białkami regulatorowymi Kalmodulina – białko wiążące wapń – jest białkiem regulatorowym Efekt działania kalmoduliny na kinazę białkową CaM

25
Działanie białek regulatorowych
Miejsce wiązania wapnia w kalmodulinie posiada charakterystyczną konformację określaną jako motyw dłoni EF. Sposób wiązania kalmoduliny z kinazą CaM

26
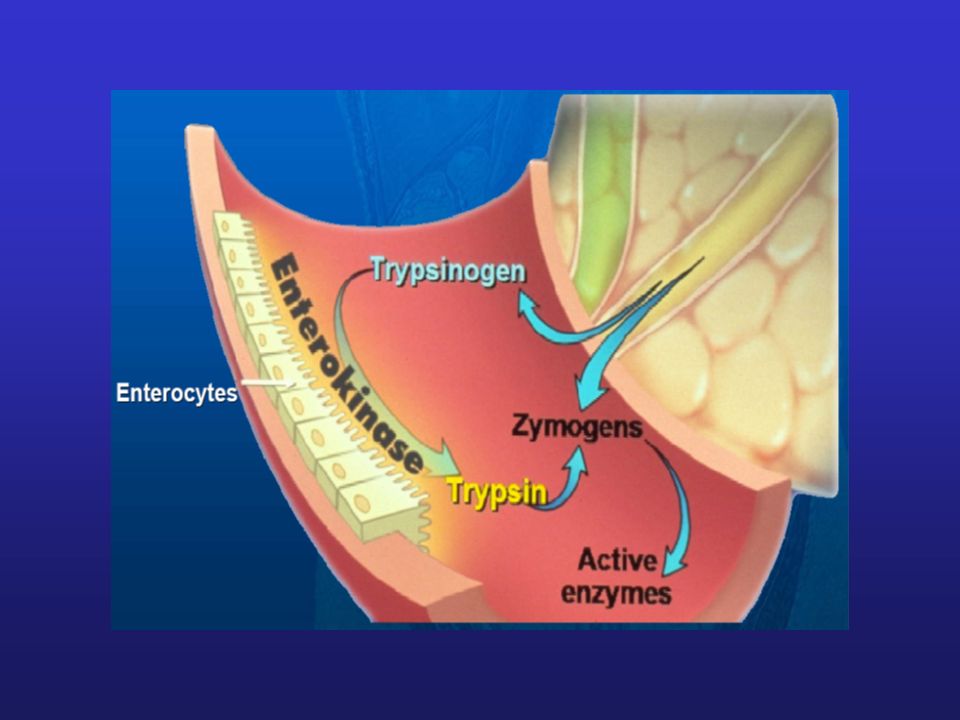
Aktywacja proteolityczna
Niektóre enzymy są syntezowane w postaci nieaktywnych zymogenów. Aktywacja zymogenów następuje w wyniku ich rozcięcia działaniem enzymów proteolitycznych. Aktywacja proteolityczna ma charakter nieodwracalny. Enzymy trawienne produkowane przez trzustkę są aktywowane proteolitycznie w jelicie cienkim Aktywacja proteolityczna chymotrypsynogenu

27
Aktywacja proteolityczna
Grupa -aminowa reszty Ile16, wygenerowana w wyniku proteolizy chymotrypsynogenu, tworzy wiązanie jonowe z Asp194 Konformacja chymotrypsynogenu i chymotrypsyny

29
Aktywacja proteolityczna
Kaskada czynników krzepliwości krwi

30
Działanie specyficznych inhibitorów
Trzustkowy inhibitor trypsyny, małe białko o masie 6 kDa, jest niezwykle skutecznym Inhibitorem trypsyny (Kd = 0.1 M). Związek ten jest analogiem substratu. działanie inhibitora zabezpiecza strukturę komórek trzustki przed konsekwencjami przedwczesnej aktywacji trypsyny.
. Związek ten jest analogiem substratu. działanie inhibitora zabezpiecza strukturę komórek trzustki przed konsekwencjami. przedwczesnej aktywacji trypsyny.")
31
Zmiany aktywności zależne od pH
Zmiany konformacyjne cząsteczki katepsyny D zachodzące w wyniku zmiany pH środowiska a/ pH obojętne – cytozol. Fragment N-końcowy blokuje centrum aktywne b/ pH < 7 – endosom. Zmiana konformacyjna powoduje odsłonięcie dostępu do centrum aktywnego

32
Autoproteoliza Niektóre białka, także enzymatyczne,
zawierają w swojej strukturze „samowycinające się” fragmenty zwane inteinami. Mechanizm autoproteolizy intein Zarówno uwolniona inteina jak i połączone eksteiny mogą wykazywać aktywność biologiczną

33
Autoproteoliza Konformacja białka zawierającego inteinę
z lewej – przed wycięciem z prawej – po wycięciu