Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
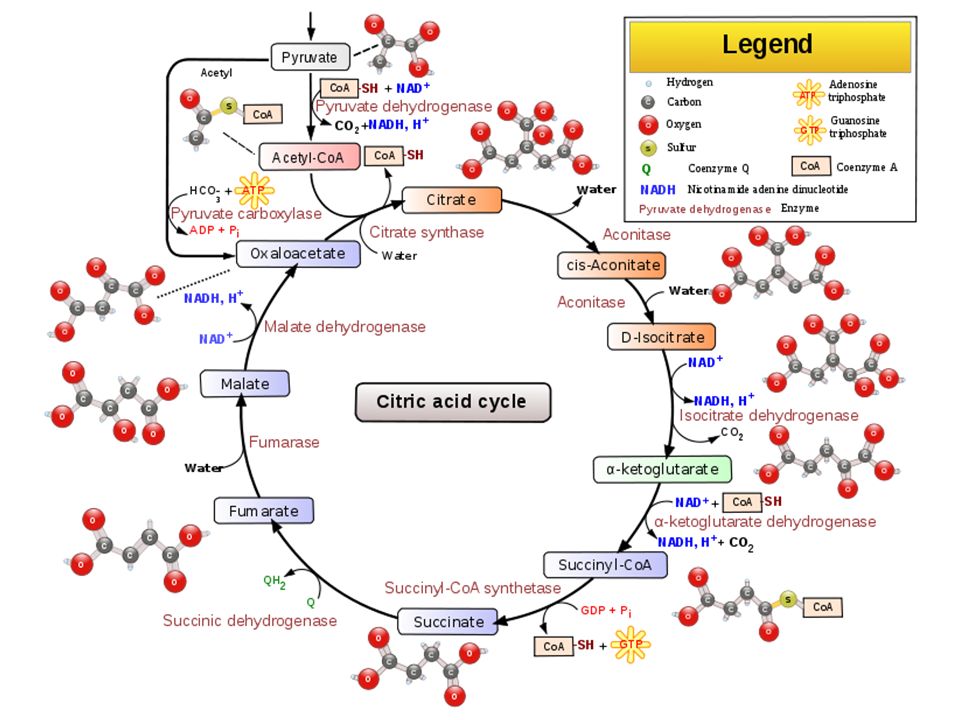
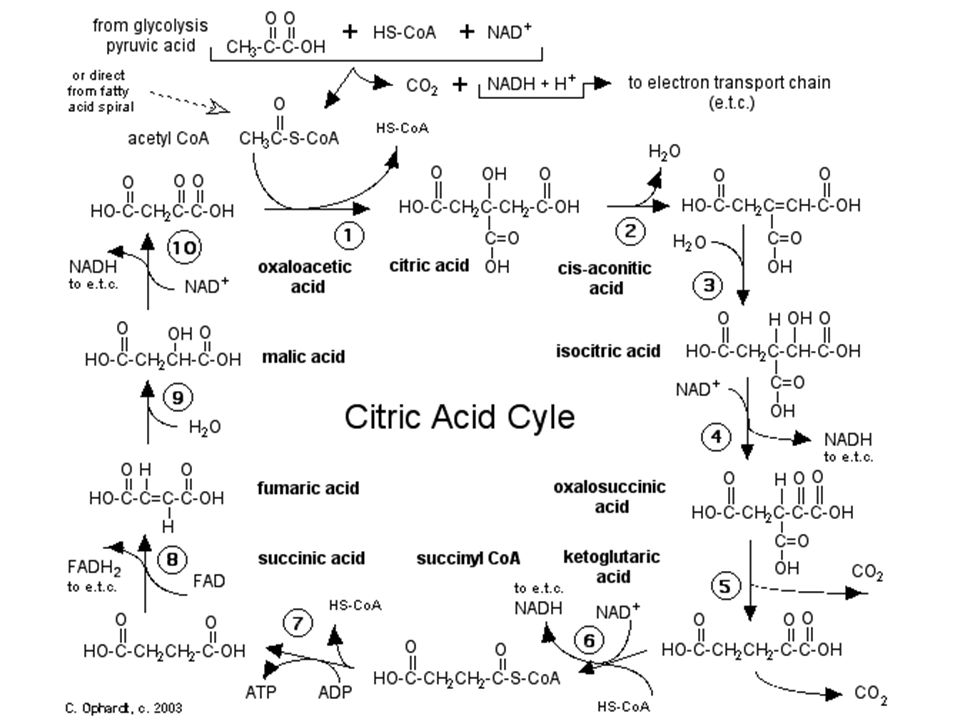
Cykl Kwasów Tri-karboksylowych = TCA = Cykl Krebsa = Cykl Kwasu Cytrynowego

2
Miejsce cyklu Krebsa na mapie metabolicznej

3
Uproszczony schemat cyklu Krebsa
(8 etapów)
")
6
Dwie drogi wejścia pirogronianu do cyklu kwasów trikarboksylowych (wątroba)
Szczawiooctan Acetylo-CoA

7
Karboksylaza pirogronianowa
Karboksylaza pirogronianowa używa kowalencyjnie przyłączonej biotyny jako kofaktora dwuetapowej reakcji karboksylacji pirogronianu. Biotyna jest najpierw łączona z CO2 pochodzącym od węglanu, (przy zużyciu ATP). Grupa karboksylowa jest następnie przenoszona przez karboksybiotynę na drugie miejsce katalityczne, gdzie pirogronian jest przekształcany w szczawiooctan.
. Grupa karboksylowa jest następnie przenoszona przez karboksybiotynę na drugie miejsce katalityczne, gdzie pirogronian jest przekształcany w szczawiooctan.")
8
Reakcja katalizowana przez dehydrogenazę pirogronianową

9
Kompleks dehydrogenazy pirogronianowej składa się z trzech białek o różnych aktywnościach. Zwróć uwagę na 5 różnych kofaktorów tego złożonego procesu. Pirogronian Acetylo-CoA

10
Kwas Liponowy pełni tu funkcję ramienia przenoszącego pozostałą po dekarboksylacji pirogronianu resztę hydroksyetylową, pomiędzy podjednostkami E1 i E3, katalizującymi kolejne etapy reakcji fragment pirgronianu Utleniony Liponian Tiamina

11
Syntaza Cytrynianowa Cytrynian powstaje w reakcji katalizowanej przez syntazę cytrynianową ze szczawiooctanu i acetylo-CoA. Mechanizm polega na ataku nukleofilowym karboanionu acetylo-CoA na węgiel karbonylowy szczawiooctanu, po czym dochodzi do hydrolizy wiązania tioestrowego.

12
Reakcje katalizowane przez Akonitazę
-H2O + H2O Kwas Cytrynowy Kwas akonitowy Kwas izocytrynowy Akonitaza (Hydrataza akonitanowa; EC ) jest enzymem katalizującym stereo-specificzną izomeryzację cytrynianu do izocytrynianu poprzez związek pośredni – kwas cis-akonitowy.
 jest enzymem katalizującym stereo-specificzną izomeryzację cytrynianu do izocytrynianu poprzez związek pośredni – kwas cis-akonitowy.")
13
Kofaktorem Akonitazy jest zespół Fe3S4

14
Akonitaza (kont.) Reakcja netto nie jest typu redoksowego!
Akonitaza zawiera zespół żelazowo-siarkowy złożony z trzech atomów żelaza i czterech atomów siarki, ułożonych w strukturę bliską kubicznej. Zespół ten jest umocowany w białku przez grupy tiolowe trzech reszt cysteiny. Brak mu jednego z narożników sześcianu. W tym właśnie miejscu wiązany jest jon F2+, odpowiedzialny za aktywację akonitazy. Ten atom koordynacyjnie wiąże C-3 karboksylu oraz grupę hydroksylową cytrynianu. Działa więc jako kwas Lewisa: przyłączając parę elektronów grupy hydroksylowej, ułatwia odłączenie całej reszty. Reakcja netto nie jest typu redoksowego!

15
Dehydrogenaza Isocytrynianowa (pierwsza reakcja redoksowa cyklu)
Dehydrogenaza Isocytrynianowa (IDH) jest enzymem cyklu pośrednio współdziałającym z łańcuchem oddechowym. Jest odpowiedzialna za odwracalne przekształcanie izocytrynianu w -ketoglutaran i CO2, w dwuetapowej reakcji. Pierwszym etapem jest utlenienie izocytrynianu do szczawiobursztynianu. W drugim etapie reakcji szczawiobursztynian traci swój -karboksyl jako CO2 .
 jest enzymem cyklu pośrednio współdziałającym z łańcuchem oddechowym. Jest odpowiedzialna za odwracalne przekształcanie izocytrynianu w -ketoglutaran i CO2, w dwuetapowej reakcji. Pierwszym etapem jest utlenienie izocytrynianu do szczawiobursztynianu. W drugim etapie reakcji szczawiobursztynian traci swój -karboksyl jako CO2 .")
16
Tlenowa dekarboksylacja -ketoglutaranu

17
Kompleks dehydrogenazy -ketoglutaranowej
-Ketoglutaran jest oksydacyjnie dekarboksylowany do bursztynylo-CoA przez dehydrogenazę -ketoglutaranową. W tej reakcji powstaje druga już cząsteczka CO2 cyklu Krebsa, oraz cząsteczka NADH. Ten kompleks wielo-enzymatyczny jest bardzo podobny do kompleksu dehydrogenazy pirogronianowej zarówno pod względem składu białkowego, rodzaju kofaktorów, jak i mechanizmu działania. Tak samo jak PDH, reakcja -KGDH przebiega z wysoką standardową zmianą energii swobodnej Gibbsa. Reakcja jest regulowana proporcjami stężeń NAD+/NADH oraz ATP/ADP.

18
Reakcja i kofaktory tlenowej dekarboksylacji -ketoglutaranu

19
Tiokinaza bursztynianowa (reakcja)
")
20
Tiokinaza bursztynylowa (Syntetaza bursztynylo - CoA)
Konwersja wysokoenergetycznego bursztynylo-CoA do wolnego bursztynianu biegnie z udziałem enzymu zdolnego do syntezy wysokoenergetycznego nukleotydu trifosforanowego. Taki proces zwiemy fosforylacją substratową. Powstaje wtedy połączenie enzym -wysokoenergetyczny związek pośredni. Dopiero z niego „wysoko energetyczna reszta fosforanowa” jest przenoszona na GDP. Powstały w ten sposób mitochondrialy GTP podlega trans-fosforylacji katalizowanej przez nukleozydo-difosfo-kinazę z cząsteczką ADP, i wytworzenie ATP. Cząsteczka GTP jest odtwarzana potem w kolejnym obrocie cyklu.

21
Dehydrogenaza Bursztynianowa (SDH)
Katalizuje utlenienie bursztynianu do fumaranu z równoczesną redukcją kowalencyjnie związanego z białkiem FAD, oraz żelaza niehemowego. W komórkach ssaków końcowym akceptorem elektronów jest koenzym Q.

22
Dehydrogenaza bursztynianowa (SDH, Kompleks II)
Skład: Cztery podjednostki białkowe (A,B,C,D) oraz następujące przenośniki elektronów: FADH2, zespoły Fe-S, hem b560, ubichinon.
 oraz następujące przenośniki elektronów: FADH2, zespoły Fe-S, hem b560, ubichinon.")
23
Fumaraza (hydrataza fumaranowa)
Katalizuje katalizuje uwodnienie fumaranu do kwasu jabłkowego.

24
Dehydrogenaza jabłczanowa (MDH)
Katalizuje odwracalną reakcję utlenienia jabłczanu do szczawiooctanu przy standardowej zmianie energii swobodnej około +7 kcal/mol. Natomiast syntaza cytrynianowa, katalizująca reakcję kondensacji acetylo-CoA ze szczawiooctanem, zachodzi przy standardowej wielkości g równej około -8 kcal/mol. Zatem to ten enzym „ciągnie” reakcję MDH w kierunku szczawiooctanu.

25
Reacja dehydrogenazy jabłczanowej

26
Sumaryczne równanie cyklu kwasów trikarboksylowych:
Acetylo-CoA + 3NAD+ + FAD + GDP + Pi + 2H2O 2CO2 + 3NADH + FADH2 + GTP + 2H+ + HSCoA

27
Energetyka cyklu Krebsa w sprzężeniu z łańcuchem oddechowym
Energetyka cyklu Krebsa w sprzężeniu z łańcuchem oddechowym. Obliczanie energii zmagazynowanej w postaci ATP W całym cyklu są: 3 utlenienia z NAD 1 utlenienie z FAD 1 fosforylacja substratowa Produkcja ATP sprzężona z tymi procesami wynosi odpowiednio: 3 x 2,5 = 7,5 cząsteczek ATP 1 x 1,5 = 1,5 cząsteczek ATP 1 x 1 = 1 GTP (= 1 cząsteczka ATP) W sumie = 10 cząsteczek ATP (utworzone z ADP) przy spaleniu jednej reszty octanowej do H2O i CO2 Jest to równoważne 310 kJ w warunkach standardowych.
 W sumie = 10 cząsteczek ATP (utworzone z ADP) przy spaleniu jednej reszty octanowej do H2O i CO2. Jest to równoważne 310 kJ w warunkach standardowych.")
28
Cykl kwasów trikarboksylowych jest przemianą amfiboliczną, czyli zarówno:
Kataboliczną jak i Anaboliczną

29
Związki z wielu przemian są katabolizowane lub przekształcane w cyklu cytrynianowym

30
Cykl cytrynianowy jako źródło prekursorów szlaków biosyntezy

31
Regulacja cyklu cytrynianowego
Regulacja tego cyklu odbywa się zarówno na wejściu jak i na poziomie samego cyklu. „Paliwo” dla tej przemiany wchodzi głównie pod postacią acetylo-CoA. Pochodzi on ze wszystkich trzech szlaków katabolicznych: lipidów, węglowodanów, aminokwasów. Jednak najistotniejszym jest źródło węglowodanowe, czyli kompleks dehydrogenazy pirogronianowej. PDH jest hamowana przez wysokie stężenia acetylo-CoA i NADH, aktywowana przez wolny CoA (CoASH) and NAD+. Mechanizm tej regulacji polega na użyciu odpowiedniej kinazy i fosfatazy. Fosforylacja PDH hamuje aktywność kompleksu, prowadząc do obniżenia tempa utleniania pirogronianu. PDH kinaza jest aktywowana przez NADH i acetyl-CoA, a hamowana przez pirogronian, ADP, CoASH, Ca2+ i Mg2+. PDH fosfataza, jest aktywowana jonami Mg2+ i Ca2+.
 and NAD+. Mechanizm tej regulacji polega na użyciu odpowiedniej kinazy i fosfatazy. Fosforylacja PDH hamuje aktywność kompleksu, prowadząc do obniżenia tempa utleniania pirogronianu. PDH kinaza jest aktywowana przez NADH i acetyl-CoA, a hamowana przez pirogronian, ADP, CoASH, Ca2+ i Mg2+. PDH fosfataza, jest aktywowana jonami Mg2+ i Ca2+.")
Podobne prezentacje





>")






, kwas siarkowy ( VI ), kwas azotowy ( V ), kwas fosforowy ( V ), kwas węglowy.>")






