Pobierz prezentację
1
Aktywność katalityczna enzymów
Enzymologia-1 Informacje wstępne Aktywność katalityczna enzymów

3
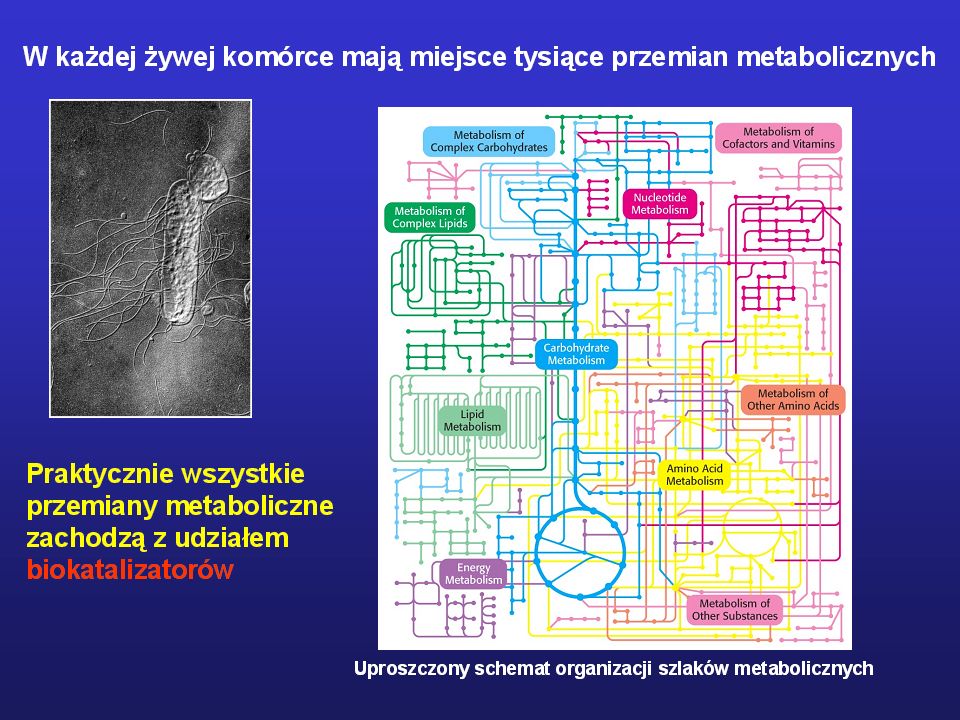
Biokatalizatory: Enzymy – zbudowane z aminokwasów (białka)
Rybozymy – zbudowane z rybonukleotydów (RNA) DNA-enzymy – zbudowane z deoksyrybonukleotydów
 DNA-enzymy – zbudowane z deoksyrybonukleotydów.")
5
Ważne daty z historii enzymologii
Kuhne – wprowadzenie terminu „enzym” 1894 Fisher – teoria „klucza i zamka” 1897 Buchner – wykazanie, że ekstrakt drożdżowy może katalizować fermentację alkoholową 1913 Michaelis, Menten – podstawy matematycznej analizy kinetyki reakcji enz, 1926 Sumner – pierwsze otrzymanie enzymu w formie krystalicznej (ureaza) 1928 Svedberg – skonstruowanie ultrawirówki 1958 Koshland – teoria „wzbudzonego dopasowania” 1960 Stein, Moore – pierwsze określenie sekwencji aminokwasowej enzymu 1965 Pierwsze określenie struktury przestrzennej enzymu (lizozym) na podstawie danych krystalograficznych. Pierwsza struktura białka (mioglobina) została określona 5 lat wcześniej (Kendrew i in.) 1965 Jacob, Monod – odkrycie zjawiska allosterycznej regulacji aktywności enzymów 1969 Merrifield – chemiczna synteza enzymu (rybonukleaza) 1970 – pierwsze wyjaśnienia mechanizmów działania enzymów 1970 Wilchek, Cuatrecas – opracowanie techniki chromatografii powinowactwa 1980 – 1985 zastosowanie technik rekombinacji DNA w badaniach enzymologicznych (układy nadekspresyjne, ukierunkowana mutageneza 1986 Cech – odkrycie rybozymu
 1928 Svedberg – skonstruowanie ultrawirówki Koshland – teoria „wzbudzonego dopasowania 1960 Stein, Moore – pierwsze określenie sekwencji aminokwasowej enzymu Pierwsze określenie struktury przestrzennej enzymu (lizozym) na podstawie. danych krystalograficznych. Pierwsza struktura białka (mioglobina) została. określona 5 lat wcześniej (Kendrew i in.) 1965 Jacob, Monod – odkrycie zjawiska allosterycznej regulacji aktywności. enzymów Merrifield – chemiczna synteza enzymu (rybonukleaza) 1970 – 1975 pierwsze wyjaśnienia mechanizmów działania enzymów Wilchek, Cuatrecas – opracowanie techniki chromatografii powinowactwa – 1985 zastosowanie technik rekombinacji DNA w badaniach. enzymologicznych (układy nadekspresyjne, ukierunkowana. mutageneza Cech – odkrycie rybozymu.")
6
Enzymy są „superkatalizatorami”
niezwykle duża efektywność katalizy wysoka aktywność katalityczna w umiarkowanych warunkach swoistość reakcji specyficzność substratowa stereospecyficzność praktycznie 100% wydajność; brak produktów ubocznych

8
SWOISTOŚĆ REAKCJI ENZYMATYCZNYCH
L-asparaginaza EC

9
SWOISTOŚĆ SUBSTRATOWA
L-asparaginaza EC

10
Stereospecyficzność działania enzymów

12
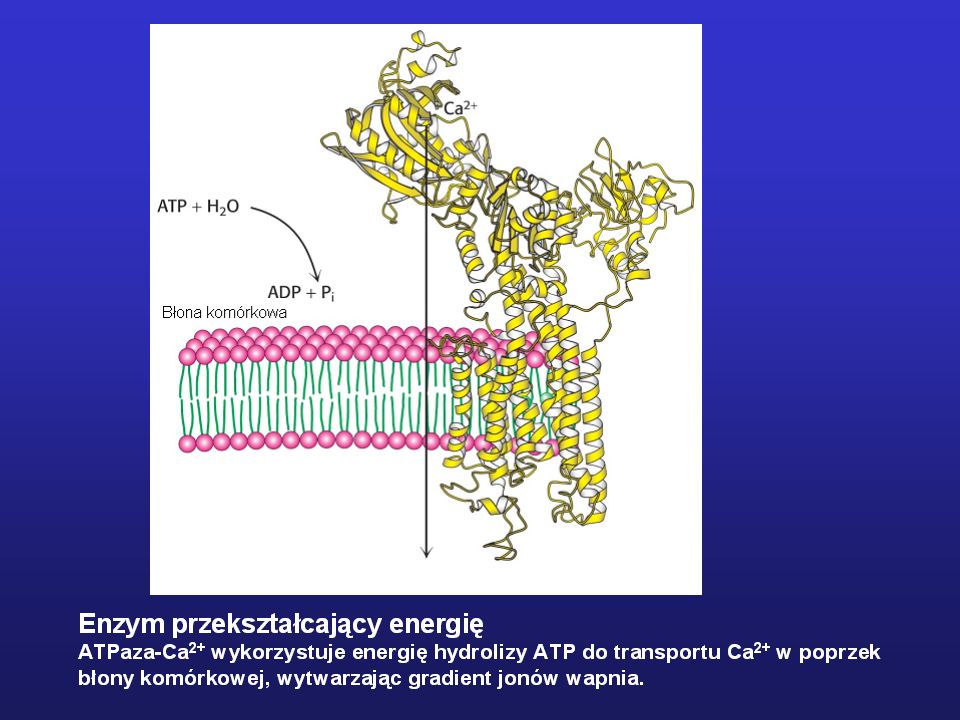
Niektóre zlokalizowane są w błonach biologicznych
Niektóre są białkami rozpuszczalnymi - globularnymi

13
Niektóre enzymy pomagają przemieszczać się substratom,
półproduktom i produktom Enzymy oligomeryczne Enzym z kanałem wewnętrznym

14
Topoizomeraza – enzym skręcający i rozkręcający DNA

15
Polimeraza DNA – enzym łączący nukleotydy
na podstawie sekwencji matrycy

17
Podobieństwo struktur przestrzennych enzymów
Struktury racemazy kwasu mandelowego I enzymu laktonizujacego kwas mukonowy Struktury dwóch proteaz serynowych: chymotrypsyny i subtilizyny

18
Dlaczego enzymy są tak efektywnymi katalizatorami?
grupy funkcyjne w centrum aktywnym hydrofobowy charakter centrum aktywnego współdziałanie koenzymów kataliza kwasowo-zasadowa maksymalne zbliżenie i optymalne ustawienie substratu(ów) wzbudzone dopasowane enzymu
 wzbudzone dopasowane enzymu.")
20
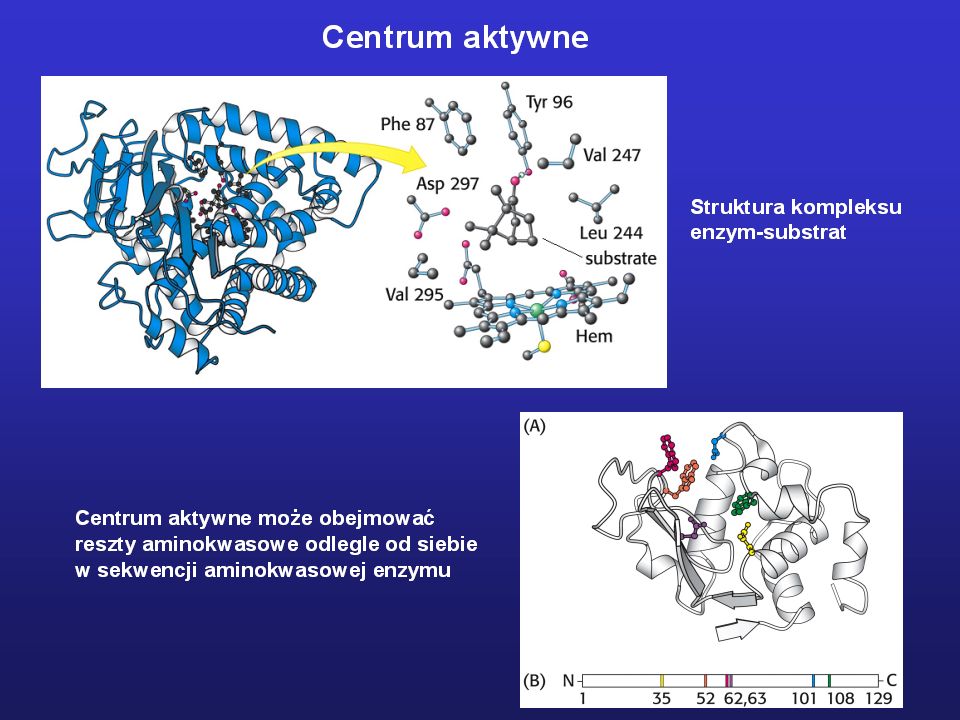
W centrum aktywnym enzymu znajdują się reszty wiążące
substrat(y) i koenzym Wiązanie koenzymu i substratu przez aminotransferazę kwasu asparaginowego Wiązanie substratu przez racemazę kwasu mandelanowego
 i koenzym. Wiązanie koenzymu i substratu. przez aminotransferazę kwasu. asparaginowego. Wiązanie substratu przez racemazę kwasu. mandelanowego.")
21
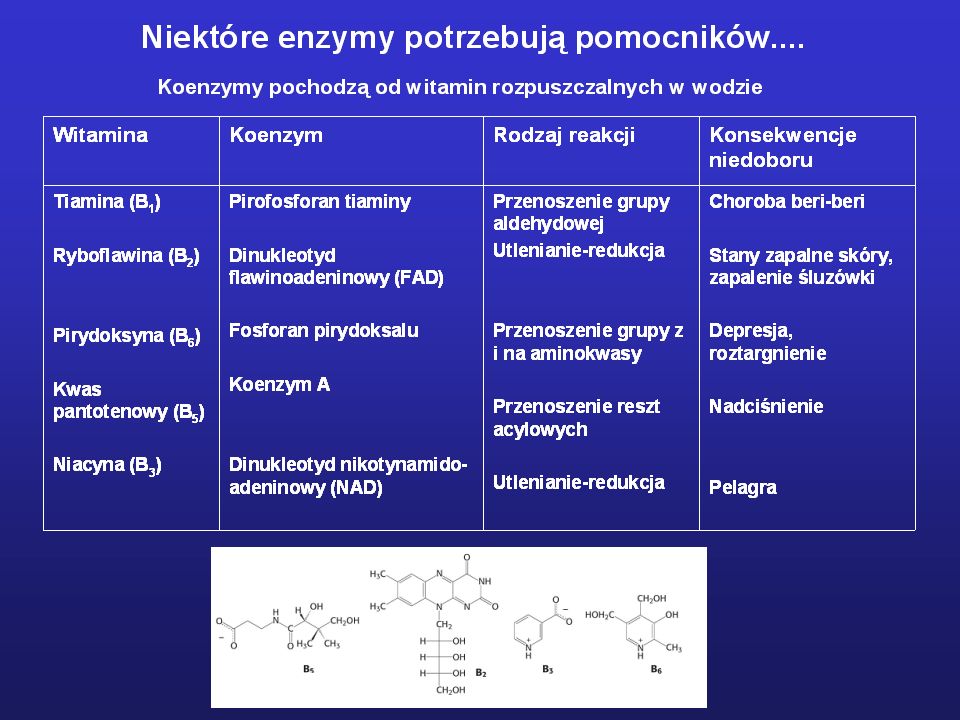
Niektóre enzymy potrzebują pomocników...
Koenzym – niewielka cząsteczka organiczna współpracująca z cząsteczką enzymu podczas aktu katalitycznego, której obecność jest niezbędna dla katalizy. Koenzym wiąże się z enzymem tylko w trakcie aktu katalitycznego Grupa prostetyczna – cząsteczka organiczna lub jon metalu niezbędna dla działania enzymu, połączona trwale z cząsteczka enzymu

23
Niektóre enzymy potrzebują pomocników...
Jony metali jako grupy prostetyczne enzymów

24
Szybkość reakcji enzymatycznej
Enzym, podobnie jak każdy inny katalizator, przyśpiesza reakcję, ale nie zmienia jej stanu równowagi. Enzymy katalizują jedynie reakcje termodynamicznie możliwe, czyli takie, dla których G0. Uwaga: enzymy w komórkach mogą katalizować reakcje, dla których G>0. Warunek: sprzężenie z reakcją, dla której G<0. Warunkiem zajścia reakcji jest efektywne zderzenie cząsteczek (cząsteczki muszą posiadać odpowiednią energię oraz być odpowiednio ustawione względem siebie) Najmniejsza energia, którą należy dostarczyć molowi substratów, aby każda z cząsteczek stała się reaktywna – energia aktywacji Szybkość reakcji zależy od energii aktywacji układu. v = f (G*)
 Najmniejsza energia, którą należy dostarczyć molowi substratów, aby każda z cząsteczek stała się reaktywna – energia aktywacji. Szybkość reakcji zależy od energii aktywacji układu. v = f (G*)")
25
Enzymy obniżają energię aktywacji układu
Energia aktywacji (kJ/mol) Połowiczny czas reakcji I rzędu w 27C 62.7 83.6 104.5 125.4 0.007 s 14 s 19 h 38 h Energia aktywacji a szybkość reakcji Energię aktywacji można wyznaczyć k = A exp(-G*/RT) ln k = f(1/T) G*
 Połowiczny czas. reakcji I rzędu w 27C s. 14 s. 19 h. 38 h. Energia aktywacji a szybkość reakcji. Energię aktywacji można wyznaczyć. k = A exp(-G*/RT) ln k = f(1/T) G*")
26
Enzymy obniżają energię aktywacji układu

27
Oddziaływanie enzym: substrat
Teoria wzbudzonego dopasowania Teoria klucza i zamka

28
Enzym wiążąc substrat przyjmuje konformację
komplementarną do stanu przejściowego

30
Enzym przyciąga substrat do centrum aktywnego
Sposoby oddziaływań elektrostatycznych enzymu z substratem Rozkład potencjału elektrostatycznego wokół enzymu – dysmutazy nadtlenkowej Pokazano obszary potencjału dodatniego i ujemnego. Substrat: O2-. jest naładowany ujemnie Efekt Kirke

31
Wykres zależności szybkości reakcji katalizowanej przez
enzym od stężenia substratu ma kształt hiperboliczny

32
Wykres zależności szybkości reakcji katalizowanej przez
enzym allosteryczny od stężenia substratu ma kształt sigmoidalny

33
Czynniki wpływające na aktywność enzymu
temperatura odczyn środowiska potencjał redoks Zależność aktywności enzymu od temperatury Zależność aktywności enzymu od pH Kształt wykresu v = f(T) może się zmieniać w zależności od momentu pomiaru
 może się zmieniać. w zależności od momentu pomiaru.")
34
Aktywność enzymów może być hamowana

35
Aktywność enzymu może być regulowana
Regulacja allosteryczna Stosunkowo niewielkie cząsteczki – ligandy allosteryczne - wiążą się z enzymami oligomerycznymi, zwiększając lub zmniejszając ich aktywność. Regulacja ma charakter płynny. Dotyczy często enzymu katalizującego pierwszą reakcję w szlaku biosyntetycznym. Regulacja kowalencyjna Enzym traci lub zyskuje aktywność w wyniku odłączenia lub przyłączenia małej grupy funkcyjnej, katalizowanego przez inny enzym. Regulacja ma najczęściej charakter zero-jedynkowy.

36
[Aktywność molekularna] = U/mol = min-1
Aktywność enzymu – ilość enzymu katalizująca przekształcenie 1 mola substratu w produkt w jednostce czasu. Jeżeli jednostką czasu jest 1 min, to aktywność jest wyrażona w jednostkach U [U] = mol/min; w układzie SI katal = mol/s 1 katal = 6 107 U Aktywność molekularna – liczba cząsteczek substratu przekształcona w produkt w czasie 1 min przez 1 cząsteczkę enzymu w optymalnych warunkach Nazwa stosowana poprzednio – liczba obrotów [Aktywność molekularna] = U/mol = min-1
![[Aktywność molekularna] = U/mol = min-1](http://slideplayer.pl/slide/417857/1/images/36/%5BAktywno%C5%9B%C4%87+molekularna%5D+%3D+U%2F%EF%81%ADmol+%3D+min-1.jpg "Aktywność enzymu – ilość enzymu katalizująca przekształcenie 1 mola. substratu w produkt w jednostce czasu. Jeżeli jednostką czasu jest 1 min, to aktywność jest wyrażona w jednostkach U. [U] = mol/min; w układzie SI katal = mol/s. 1 katal = 6 107 U. Aktywność molekularna – liczba cząsteczek substratu przekształcona. w produkt w czasie 1 min przez 1 cząsteczkę enzymu w optymalnych. warunkach Nazwa stosowana poprzednio – liczba obrotów. [Aktywność molekularna] = U/mol = min-1.")
37
Aktywność molekularna 60 = kkat
Stała katalityczna Stała katalityczna określa liczbę mikromoli substratu przekształconych przez 1 mol enzymu w produkt w ciągu 1 sekundy w optymalnych warunkach kkat = V/[E] [kkat] = s-1 Aktywność molekularna 60 = kkat Dla enzymów zawierających więcej niż jedno centrum katalityczne Aktywność centrum katalit. = aktywność molekularna/liczba centrów

38
Izoenzymy Różne formy tego samego enzymu występujące w różnych organelach komórki lub różnych tkankach tego samego organizmu. Izoenzymy mogą się różnić m.in. strukturą przestrzenną, stopniem ufosforylowania, sposobem regulacji aktywności. Różnice te są konsekwencją niewielkich zmian struktury I-rzędowej.

40
Autokatalityczne introny Rybozymy typu „głowa młotka” i enzymy DNA
Rybozymy i enzymy DNA Cząsteczki RNA lub DNA wykazujące aktywność autokatalityczną lub katalityczną RNaza P Autokatalityczne introny Rybozymy typu „głowa młotka” i enzymy DNA Transferaza peptydylowa w rybosomie

41
Autokatalityczne introny
Struktura autokatalitycznego intronu z prekursora rRNA Tetrahymena, wycinającego się bez pomocy białek Mechanizm autokatalitycznego splicingu

42
Niektóre rybozymy i enzymy DNA przecinają mRNA
Modele struktur II-rzędowych rybozymu typu „głowa młotka” (ang. hammerhead) i enzymu DNA Konformacja zwiniętego kompleksu rybozymu typu „głowa młotka” z mRNA
 i enzymu DNA. Konformacja zwiniętego kompleksu. rybozymu typu „głowa młotka z mRNA.")
43
Transferaza peptydylowa – największy rybozym

44
Rybosom

45
Obraz miejsca tworzenia wiązania peptydowego w rybosomie


















 , chlorki , siarczki, azotany (V), węglany, fosforany (V), siarczany (IV).>")






>")
