Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
Metody określania struktury enzymów (część II)
Enzymologia-4 Metody określania struktury enzymów (część II)
")
2
2. Określanie struktury III- i IV-rzędowej
2.1 Krystalografia i analiza rentgenograficzna białek - krystalizacja białek - aparatura rentgenograficzna - otrzymywanie danych i ich interpretacja 2.2 NMR w badaniu struktury przestrzennej białek 2.3 Modelowanie molekularne 2.4 Badania spektralne 2.5 Określanie struktury IV-rzędowej

3
Struktura trzeciorzędowa to przestrzenne ułożenie całego polipeptydu, czyli jego kształt, który jest stabilizowany wzajemnymi oddziaływaniami bocznych reszt aminokwasowych.

5
Specyficzne cechy kryształów białek
niewielkie rozmiary; zwykle < 1mm; zawierają zwykle niewiele elementów symetrii, a w konsekwecji wykazują dość proste kształty; kryształy białek są szczególnie wrażliwe i niestabilne; wynika to z dużej zawartości rozpuszczalnika (zwykle 40 – 70%); kryształy białek są wrażliwe na zmiany pH, siły jonowej, temperatury; stabilność w niskich temperaturach można poprawić poprze użycie krioprotektantów; kryształy białek często słabo rozpraszają promieniowanie rentgenowskie, z rozdzielczością niezadowalającą dla otrzymania danych strukturalnych
; kryształy białek są wrażliwe na zmiany pH, siły jonowej, temperatury; stabilność. w niskich temperaturach można poprawić poprze użycie krioprotektantów; kryształy białek często słabo rozpraszają promieniowanie rentgenowskie, z rozdzielczością niezadowalającą dla otrzymania danych strukturalnych.")
6
Składniki roztworów używanych do krystalizacji białek
i warunki krystalizacji Czynniki wspomagające tworzenie kryształów: siarczan amonu, siarczan litu, siarczan magnezu, siarczan sodu, cytrynian sodu, mrówczan sodu, glikol polietylenowy , 2-metylopentandiol, alkohol polywinylowy, dekstran Stabilizatory: EDTA, ditiotreitol, inhibitory proteaz, detergenty niejonowe (redukują agregację cząsteczek białka) Krioprotektant: glicerol Warunki: temperatura (zwykle 4 C, 15 C, pokojowa), pH (raczej nie pH = pI) Uwaga: warunki sprzyjające tworzeniu zarodków kryształów są często różne od tych, które sprzyjają wzrostowi kryształów
 Krioprotektant: glicerol. Warunki: temperatura (zwykle 4 C, 15 C, pokojowa), pH (raczej nie pH = pI) Uwaga: warunki sprzyjające tworzeniu zarodków kryształów są często różne od tych, które sprzyjają. wzrostowi kryształów.")
7
Otrzymywanie kryształów kompleksów enzym:ligand
współkrystalizacja enzymu z ligandem nasączanie kryształów enzymu stężonym roztworem liganda Enzym – jeden z substratów Enzym – inhibitor allosteryczny Enzym – inhibitor, analog stanu przejściowego

8
Optymalizacja warunków krystalizacji
Testy przesiewowe Metoda kropli siedzącej Metoda kropli wiszącej Mikropłytka stosowana do badań przesiewowych

9
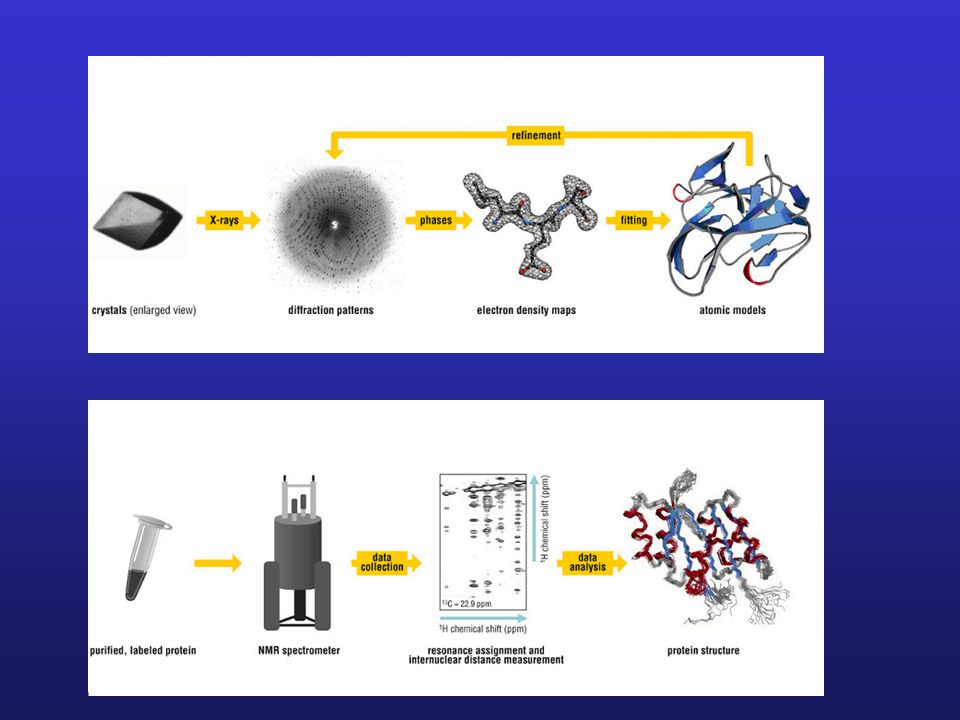
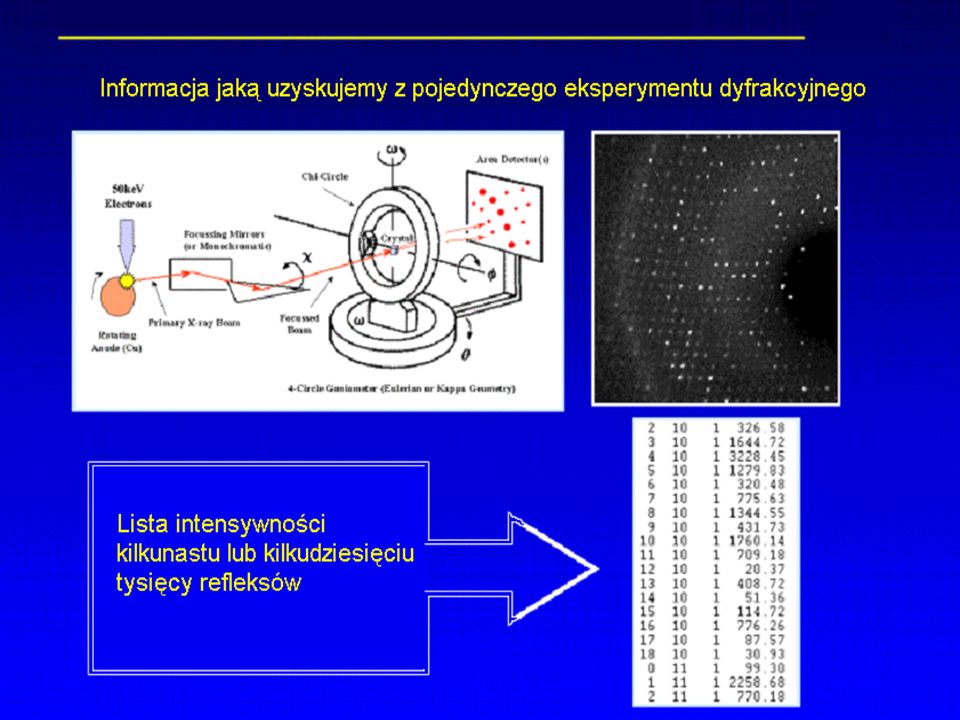
Aparatura do rentgenografii strukturalnej
Dyfraktometr

10
Wysokoenergetyczne elektrony są przyśpieszane w akceleratorze kołowym.
Kształt orbity biegu elektronów jest kontrolowany przez układ magnesów. Akcelerator o promieniu 200 m umożliwia uzyskanie energii 100 GeV. Wygenerowany strumień promieni X jest wąski i ekstremalnie intensywny (~100 x ). Lokalizacja synchrotronów: USA Cornell, Stanford, Argonne, Los Alamos UK Daresbury France Grenoble (EMBL) Germany Hamburg (EMBL) Japan Photon Factory Synchrotron w Grenoble
. Lokalizacja synchrotronów: USA. Cornell, Stanford, Argonne, Los Alamos. UK. Daresbury. France. Grenoble (EMBL) Germany. Hamburg (EMBL) Japan. Photon Factory. Synchrotron w Grenoble.")
11
W klasycznym układzie pomiarowym
kryształ jest obracany powoli, prostopadle do monochromatycznej wiązki promieni X. Kryształ umieszczony na pętli włókna Kryształ białka w równowadze z ciekłym rozpuszczalnikiem w cienkościennej kapilarze.

12
Detektory promieniowania X
Błona fotograficzna Charge-coupled device (CCD) Płyta Detektor FAST
 Płyta. Detektor FAST.")
14
(a) Mapa o niskiej rozdzielczości (5 Å lub więcej – ogólny kształt cząsteczki;
(b) Mapa o średniej rozdzielczości (~3 Å) – przebieg łańcucha polipeptydowego możliwość dopasowania znanej sekwencji aminokwasowej do mapy; (c) Mapa o wysokiej rozdzielczosci (1.5 - Å) – jednoznaczna identyfikacja reszt aminokwasowych; (d) Mapa o bardzo wysokiej (atomowej) rozdzielczości (poniżej 1 Å) – atomy jako izolowane kuleczki gęstości elektronowej Rozdzielczość danych dyfrakcyjnych zależy od wielkości kryształu i stopnia jego uporządkowania
 Mapa o średniej rozdzielczości (~3 Å) – przebieg łańcucha polipeptydowego. możliwość dopasowania znanej sekwencji aminokwasowej do mapy; (c) Mapa o wysokiej rozdzielczosci (1.5 - Å) – jednoznaczna identyfikacja reszt. aminokwasowych; (d) Mapa o bardzo wysokiej (atomowej) rozdzielczości (poniżej 1 Å) – atomy jako. izolowane kuleczki gęstości elektronowej. Rozdzielczość danych dyfrakcyjnych zależy od wielkości kryształu i stopnia jego. uporządkowania.")
15
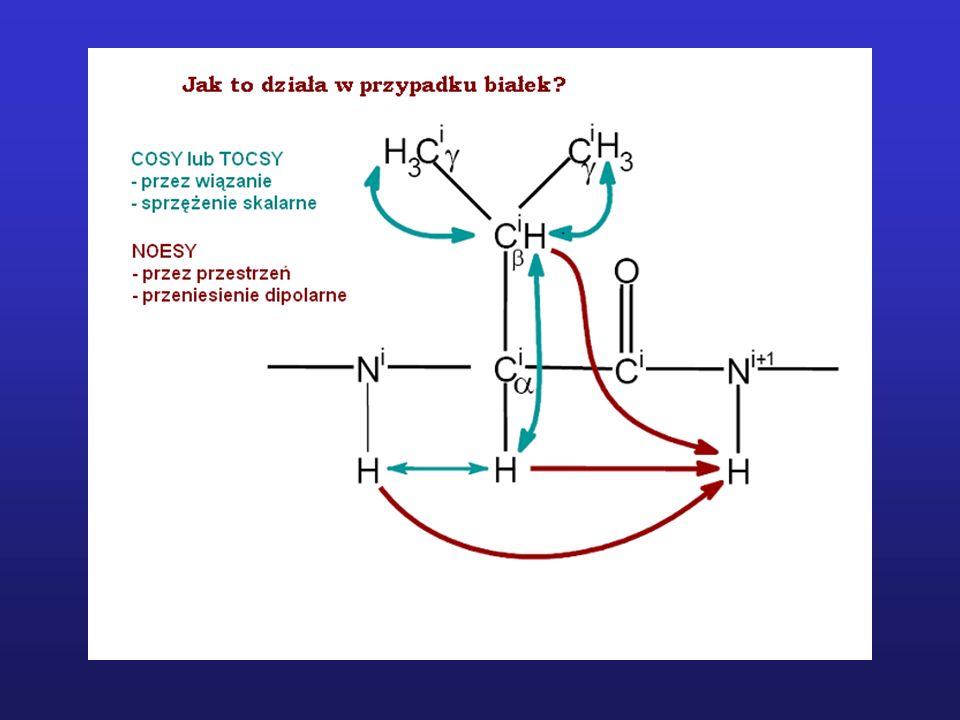
Zastosowanie NMR do badania struktury białek
Widmo 1H NMR niewielkiego białka (< 15 kDa)
")
16
Spektroskopia NMR Schemat budowy spektrometru NMR
Podstawa metody: zjawisko magnetyzmu jądrowego Zasada metody: Znajdujące się w zewnętrznym polu magnetycznym jądra atomów posiadające moment magnetyczny (np. 1H, 19F, 14N, 15N, 13C), na które działa promieniowanie elektromagnetyczne o częstotliwości radiowej, absorbują kwanty energii tego promieniowania, przechodząc na wyższy poziom energetyczny. Absorpcję tą określa się jako jądrowy rezonans magnetyczny. Absorpcja energii uwidacznia się jako układ linii spektralnych – sygnałów rezonansowych. Schemat budowy spektrometru NMR
, na które działa promieniowanie. elektromagnetyczne o częstotliwości radiowej, absorbują kwanty energii tego. promieniowania, przechodząc na wyższy poziom energetyczny. Absorpcję tą. określa się jako jądrowy rezonans magnetyczny. Absorpcja energii uwidacznia się jako. układ linii spektralnych – sygnałów. rezonansowych. Schemat budowy spektrometru NMR.")
18
Wzrost ilości rozwiązanych struktur białek i kwasów nukleinowych w latach 1972-2004

19
Modelowanie molekularne
Określenie struktury trzeciorzędowej białka na podstawie sekwencji aminokwasowej jest możliwe pod warunkiem, iż znana jest struktura jednego lub kilku białek homologicznych Metoda opiera się na założeniu, że białka homologiczne tworzą podobną strukturę przestrzenną. Dostępne programy, stosujące takie podejście to np.: COMPOSER oraz SWISS-MODEL

20
Model domeny glutaminowej syntazy GlcN-6-P z C
Model domeny glutaminowej syntazy GlcN-6-P z C. albicans (kolor żółty), struktura domeny glutaminowej syntazy GlcN-6-P z E. coli (kolor ciemnoróżowy)
, struktura domeny glutaminowej syntazy GlcN-6-P z E. coli (kolor ciemnoróżowy)")
21
Badania spektralne Zastosowanie metod spektralnych, często obok innych metod fizykochemicznych i chemicznych, w wielu przypadkach pozwala na uzyskanie dokładnych informacji o strukturze enzymów i przekształceniu ich konformacji przy przyłączeniu koenzymów, substratów lub inhibitorów.

22
Metoda różnicowej spektroskopii absorpcyjnej
Pochłanianie w obszarze nm (daleki ultrafiolet) jest wynikiem elektronowego wzbudzenia atomów w wiązaniach peptydowych, a w obszarze nm (bliski ultrafiolet) jest wynikiem wzbudzenia atomów w pierścieniach reszt Tyr, Trp oraz w mniejszym stopniu Phe. absorpcja (nm) Phe 260 Tyr 275 Trp 280
 jest wynikiem elektronowego wzbudzenia atomów w wiązaniach peptydowych, a w obszarze nm (bliski ultrafiolet) jest wynikiem wzbudzenia atomów w pierścieniach reszt Tyr, Trp oraz w mniejszym stopniu Phe. absorpcja. (nm) Phe Tyr Trp")
23
Zmiany widm pochłaniania
Widmo pochłaniania promieniowania przez chromofory zależy od tego czy znajdują się one w środowisku cząsteczek wody, a więc w środowisku o wysokiej stałej dielektrycznej, czy też w środowisku bezwodnym - o niskiej stałej dielektrycznej. Obniżenie stałej dielektrycznej mikrootoczenia chromoforu powoduje przesunięcie widma w kierunku fal dłuższych tzw.: przesunięcie batochromowe lub „czerwone”. Zwiększenie stałej dielektrycznej mikrootoczenia chromoforu (najczęściej podczas rozfałdowywania białka) powoduje przesunięcie widma w kierunku fal krótszych tzw.: przesunięcie hypsochromowe lub „niebieskie”.
 powoduje przesunięcie widma w kierunku fal krótszych tzw.: przesunięcie hypsochromowe lub „niebieskie .")
24
Przykład: Próbka enzymu w buforze Próbka enzymu w buforze z dodatkiem 6M mocznika Wyniki: Zmiany w widmie – reszty Trp usytuowane wewnątrz struktury enzymu, po denaturacji zmiany charakteru mikrośrodowiska reszt Trp Brak zmian w widmie – reszty Trp usytuowane na powierzchni W przypadku przesunięcia „czerwonego” powstaje dodatnie widmo różnicowe, a przesunięcia „niebieskiego” - ujemne widmo różnicowe

25
Perturbacja (zaburzanie) widm
Czynnik perturbujący - dodany do próbki enzymu nie powoduje denaturacji białka, ale wywołuje zmiany w jego widmie sacharoza glicerol acetonitryl dimetyloformamid

26
Widmo różnicowe fosfoglukomutazy powstające przy związani substratu – glukozo-6-fosforanu

27
Spektrofluorymetria Metoda, która wykorzystuje zdolność reszt Trp, Tyr, Phe do emisji części zaabsorbowanej energii w formie promieniowania elektromagnetycznego w zakresie UV lub w obszarze widzialnym. Proces ten nazywa się fluorescencją. absorpcja (nm) emisja Phe 260 282 Tyr 275 303 Trp 280 348 Widmo emisji ( zielone) i absorpcji ( niebieskie) Poziomy energetyczne w cząsteczce, S1 – stan podstawowy, S0 – stan wzbudzenia
 emisja. Phe Tyr Trp Widmo emisji ( zielone) i absorpcji ( niebieskie) Poziomy energetyczne w cząsteczce, S1 – stan podstawowy, S0 – stan wzbudzenia.")
28
Różnice w długościach fal dla maksymalnej emisji fluorescencji wymienionych aminokwasów są znacznie większe aniżeli różnice w poszczególnych maksimach absorpcji, co oznacza, że pomiary fluorescencji są bardziej selektywne, niż pomiary absorpcji. W białkach zawierających wszystkie trzy aminokwasy obserwowana fluorescencja pochodzi z reszt tryptofanylowych, z powodu niskiej wydajności kwantowej Tyr i jeszcze niższej Phe.

29
zmiana środowiska chromoforu na bardziej hydrofobowe powoduje wzrost intensywności fluorescencji i przesuwanie maksimum w kierunku fal krótszych, zmiana środowiska chromoforu na bardziej hydrofilowe powoduje obniżenie intensywności fluorescencji i przesuwanie maksimum w kierunku fal dłuższych.

30
Zwiększenie wrażliwości metody – dodatek grup sprawozdawczych
1-anilinonaftaleno-8-sulfonian (ANS) W roztworach wodnych nie posiada właściwości fluorescencyjnych, natomiast po niekowalencyjnym połączeniu się z częścią hydrofobową białka powoduje kilkusetkrotne zwiększenie fluorescencji białka
 W roztworach wodnych nie posiada właściwości fluorescencyjnych, natomiast po niekowalencyjnym połączeniu się z częścią hydrofobową białka powoduje kilkusetkrotne zwiększenie fluorescencji białka.")
31
Zmiany widm spektrofluorymetrycznych zmutowanej amidotransferazy PRPP (mutacje C1S i W290F) pod wpływem związania substratów: L-Gln i/lub PRPP
 pod wpływem związania substratów: L-Gln i/lub PRPP")
32
Spektropolarymetria W metodach spektropolarymetrycznych stosuje się światło spolaryzowane Kiedy światło spolaryzowane w płaszczyźnie przechodzi przez związki o budowie asymetrycznej obserwuje się tzw. Efekt Cottona. Ugrupowania boczne białek globularnych wywołują efekt Cottona zwany Efektem Cottona łańcuchów bocznych w obszarze nm.

33
Efekt jest wynikiem zakłóconej rotacji atomów węgli a-Cys, Phe, Trp i Tyr w białkach.
Dodatnie widmo CD przy nm jest wynikiem pochłaniania światła przez mostki -S-S- , ujemne zaś w obszarze nm pochłaniania przez aromatyczne ugrupowanie Tyr i reszty Trp. Widmo dichroizmu kołowego wolnej Co-karboksypeptydazy oraz związanej z substratem lub inhibitorem. a – Co-karboksypeptydaza, b - Co-karboksypeptydaza + glicylo-L-tyrozyna (substrat), c - Co-karboksypeptydaza + b-fenylopropionian (inhibitor).
, c - Co-karboksypeptydaza + b-fenylopropionian (inhibitor).")
34
Określanie czwartorzędowej struktury białka
Porównanie wielkości MW białka natywnego (chromatografia rozmiarów wykluczających, ultrawirowanie) oraz zdenaturowanego (SDS-PAGE ) 2. Sieciowanie białka Sieciowane białek w wyniku działania diestru metylowego kwasu suberowego sieciowanie; b. analiza SDS-PAGE 3. Określanie stechiometrii wiązania ligandów
 oraz zdenaturowanego. (SDS-PAGE ) 2. Sieciowanie białka. Sieciowane białek w wyniku działania diestru metylowego kwasu suberowego. sieciowanie; b. analiza SDS-PAGE. 3. Określanie stechiometrii wiązania ligandów.")
35
Sieciowanie białek + Lys
Bis(Sulfosuccinimidyl)suberate, Długość ramienia 11.4Å N SO 3 Na NaO S Protein Lys N H 2 + O N H Białko usieciowane
suberate, Długość ramienia 11.4Å. N. SO. 3. Na. NaO. S. Protein. Lys. N. H O. N. H. Białko usieciowane.")
Podobne prezentacje














>")




