Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
油菜育种讲义 油菜育种讲义 一、油菜育种概况 最早:油菜进行育种最早的是德国人 Hans Lembke , 他于 1897 年开始油菜育种, 1910 年他首次建立了 油菜科学育种体系,即个体选择和系谱测验, 1917 年育成第一个高产的甘蓝型冬性品种 “Lembke Winterraps” ,该品种在德国和欧洲各国作为高产品 种应用长达半个世纪。 德国、日本、瑞典:开展油菜育种比较早。 加拿大、澳大利亚等 :出口国家 中国:进口国

3
重要的进展 其一,油菜的亲缘关系与实验进化育种 禹长春等( 1935 ); Olsson,G 等 其二,油菜的优质资源的发现和利用 加拿大 Stefanson 等( 50 年代) Krzymanski,J. ( 1968 ) 其三,雄性不育资源发现和杂种优势利用 Polima cms 、 nap cms 、陕 2A 等 核不育 其四,生物技术在油菜研究上的广泛应用 抗除草剂、雄性 MS 、细胞融合、小孢子培养
 其三,雄性不育资源发现和杂种优势利用 Polima cms 、 nap cms 、陕 2A 等 核不育 其四,生物技术在油菜研究上的广泛应用 抗除草剂、雄性 MS 、细胞融合、小孢子培养.")
4
二、油菜常规品种选育 育种目标: 油菜的高产育种 ; 双低油菜育种 ; 高含油量育种 ; 抗逆性品种选育 ; 无花瓣性状 、单倍体 、实验进化 、 特异脂肪酸构成

5
高产育种 遗传上,产量相关因素的遗传率大小次 序为:果长 > 开花期 > 千粒重 > 主花序长度 > 主花序结角密度 > 每角粒数 > 着角密度 > 分枝部位 > 株高 > 主花序角果数 > 单株产量 > 总角果数(刘定富, 1984 )。 要求:各个产量构成要素综合协调 综合遗传学、生理知识,指导育种过程。
。 要求:各个产量构成要素综合协调 综合遗传学、生理知识,指导育种过程。")
6
双低油菜育种 ,芥酸的碳链比较长,食用后易在血管 中积累胆固醇等物质,可能造成心脑血 管疾病。降低芥酸含量,则油酸等短链、 不饱和脂肪酸含量则会提高,这些短链 脂肪酸有益利于人体,因而减低芥酸含 量会提高菜油的利用价值。 食用菜油一般要求芥酸含量小于 1% ,商 品菜籽收购时国际标准采用小于 2% 。

7
脂肪酸构成 菜油中,主要有硬脂酸 (Stearic acid) 、油酸 (Oleic acid) 、亚油酸 (Linoeic acid) 、二十烷烯酸 (Eicosenoic acid) 、芥酸( Erucic acid )、亚麻酸 (Linoleic acid) 等。脂肪酸的生物合成主要是通过 油酸向两个方面进行的,一是碳链的减饱和作用, 以油酸( C 18 : 1 )为前体,增加不饱和度生成亚油 酸( C 18 : 2 ),再生成亚麻酸( C 18 : 3 );二是通 过碳链的延长作用,把一个乙酸分子加到油酸的 羧基末端,组成二十烷烯酸( C 20 : 1 ),再加上一 个乙酸分子组成芥酸( C 22 : 1 )。
 、油酸 (Oleic acid) 、亚油酸 (Linoeic acid) 、二十烷烯酸 (Eicosenoic acid) 、芥酸( Erucic acid )、亚麻酸 (Linoleic acid) 等。脂肪酸的生物合成主要是通过 油酸向两个方面进行的,一是碳链的减饱和作用, 以油酸( C 18 : 1 )为前体,增加不饱和度生成亚油 酸( C 18 : 2 ),再生成亚麻酸( C 18 : 3 );二是通 过碳链的延长作用,把一个乙酸分子加到油酸的 羧基末端,组成二十烷烯酸( C 20 : 1 ),再加上一 个乙酸分子组成芥酸( C 22 : 1 )。")
8
芥酸的遗传 芥酸含量受胚基因型控制,由两对具有 加性效应的基因控制 (Harvey,B.L. 等, Downey,R.K.,1964) 。白菜型油菜的芥酸 含量受 A 染色体组上的一个位点控制。芥 菜型和埃塞俄比亚芥的芥酸含量都受两 个位点控制。据 Downey 等研究,甘蓝型 油菜控制芥酸的 2 个位点的 4 个基因各控 制芥酸含量 9~10% 。
 。白菜型油菜的芥酸 含量受 A 染色体组上的一个位点控制。芥 菜型和埃塞俄比亚芥的芥酸含量都受两 个位点控制。据 Downey 等研究,甘蓝型 油菜控制芥酸的 2 个位点的 4 个基因各控 制芥酸含量 9~10% 。.")
9
硫甙 菜籽饼粕中,含有硫甙,白菜型含量为 3% ,甘 蓝型含量为 4~7% ,芥菜型含量为 5~7% 。硫甙在 芥子酶或水解酶的作用下,可以分解为恶唑烷硫 酮、异硫氰酸盐和睛等有毒物质,可使单胃动物 的甲状腺肿大,并导致代谢混乱,但对反刍动物 无害。低硫甙品种的菜饼可饲喂猪、鸡。 甘蓝型油菜中,硫甙水解产物为恶唑烷硫酮、异 硫氰酸盐。脱脂后饼粕中异硫氰酸盐降低到 0.5~ 1.5 毫克 / 克,才不致使家畜中毒。 育种要求降低硫甙含量到小于 40 微摩尔 / 克(含 吲哚硫甙),或小于 30 微摩尔 / 克(不含吲哚硫 甙),欧洲的硫甙标准近年来有所提高,要求硫 甙含量小于 10~20 微摩尔 / 克。
,或小于 30 微摩尔 / 克(不含吲哚硫 甙),欧洲的硫甙标准近年来有所提高,要求硫 甙含量小于 10~20 微摩尔 / 克。")
10
硫甙遗传 遗传上硫甙是 3 对或 3 对以上基因控制的 ( Morice,J. 1980 ),低硫甙性状在遗传 上为隐性,但是不同类型的油菜的硫甙 成分不同,含量也因为品种而异。低硫 甙性状往往与不良农艺性状间存在连锁, 但是低硫甙与不良农艺性状的连锁可以 通过长期育种过程来打破的。
,低硫甙性状在遗传 上为隐性,但是不同类型的油菜的硫甙 成分不同,含量也因为品种而异。低硫 甙性状往往与不良农艺性状间存在连锁, 但是低硫甙与不良农艺性状的连锁可以 通过长期育种过程来打破的。.")
11
品质:播娘蒿硫苷成分 R 基

12
高含油量育种 多世代的单株选择 种间杂交 黄籽油菜育种 刘后利( 1975 )发现了甘蓝型油菜的黄籽突变体, 遗传研究表明是受 3 对隐性基因控制,且发现在 同等遗传背景下的含油量高 1.2~4.3% ,还育成了 黄籽品种华黄 1 号,其含油量比普通黑籽品种高 5~8% 。 吴江生等( 1998 )又发现了两个位点控制的显性 黄籽种质,黄籽基因型为 YcYcbcbc , Bc 基因的 出现可以抑制黄籽的产生,形成黑籽。
发现了甘蓝型油菜的黄籽突变体, 遗传研究表明是受 3 对隐性基因控制,且发现在 同等遗传背景下的含油量高 1.2~4.3% ,还育成了 黄籽品种华黄 1 号,其含油量比普通黑籽品种高 5~8% 。 吴江生等( 1998 )又发现了两个位点控制的显性 黄籽种质,黄籽基因型为 YcYcbcbc , Bc 基因的 出现可以抑制黄籽的产生,形成黑籽。")
13
不同含油量的资源

15
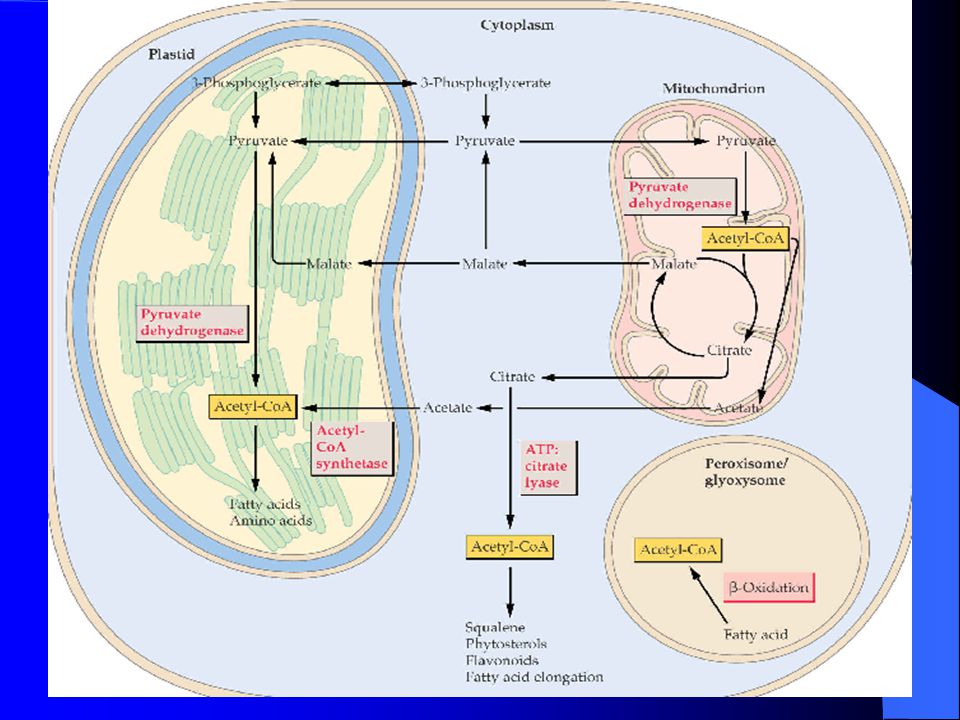
油菜油脂合成

16
脂肪酸合成

17
抗逆性品种选育 抗逆性包括:抗病虫性、抗寒性、耐旱 性和耐湿性、耐盐性、耐瘠性等。 甘蓝型抗性较强,芥菜型次之,白菜型 感病一般较严重。油菜的病害还包括黑 胫病、根肿病、白锈病等,这些病害在 我国发病不普遍。 抗裂角性 、抗倒伏等 转基因:抗除草剂、抗病、抗虫

18
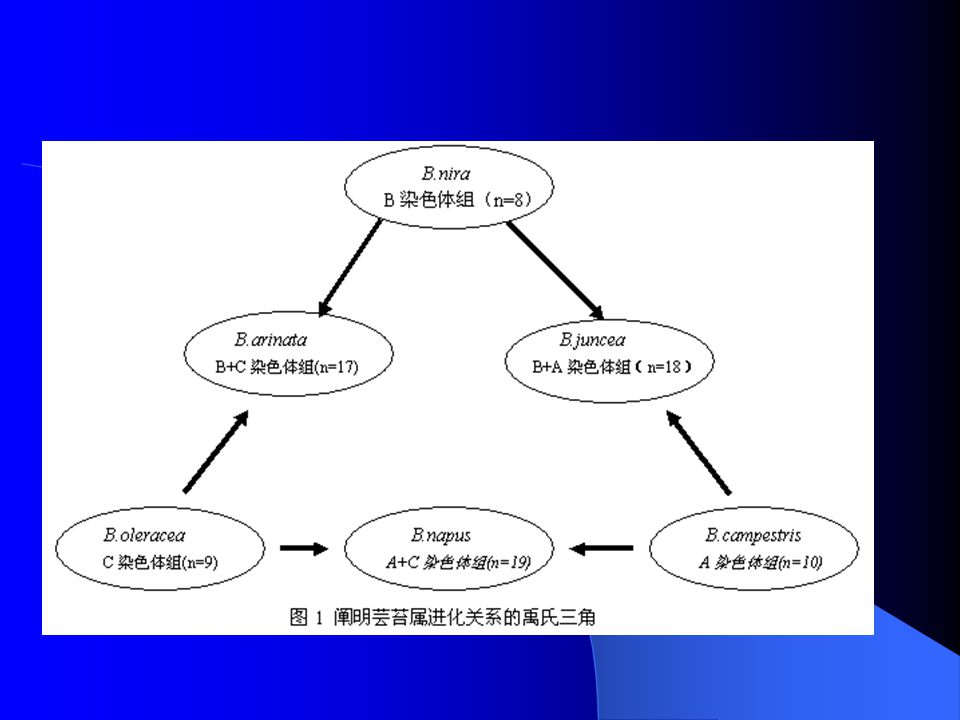
油菜常规育种的其它方面 无花瓣品种 : 3 对隐性基因。抗病、提高 物质利用效率 单倍体育种 :自 1982 年 Lichter 等用甘蓝型 油菜游离小孢子获得再生植株以来,油菜 小孢子培养技术逐步成熟 油菜实验进化育种 :白菜与甘蓝的种间杂 交人工合成甘蓝型油菜 特异脂肪酸构成的品种选育 :任意脂肪酸 含量高都能够大大提高效益。

19
油菜与播娘蒿融合

20
花和花粉

21
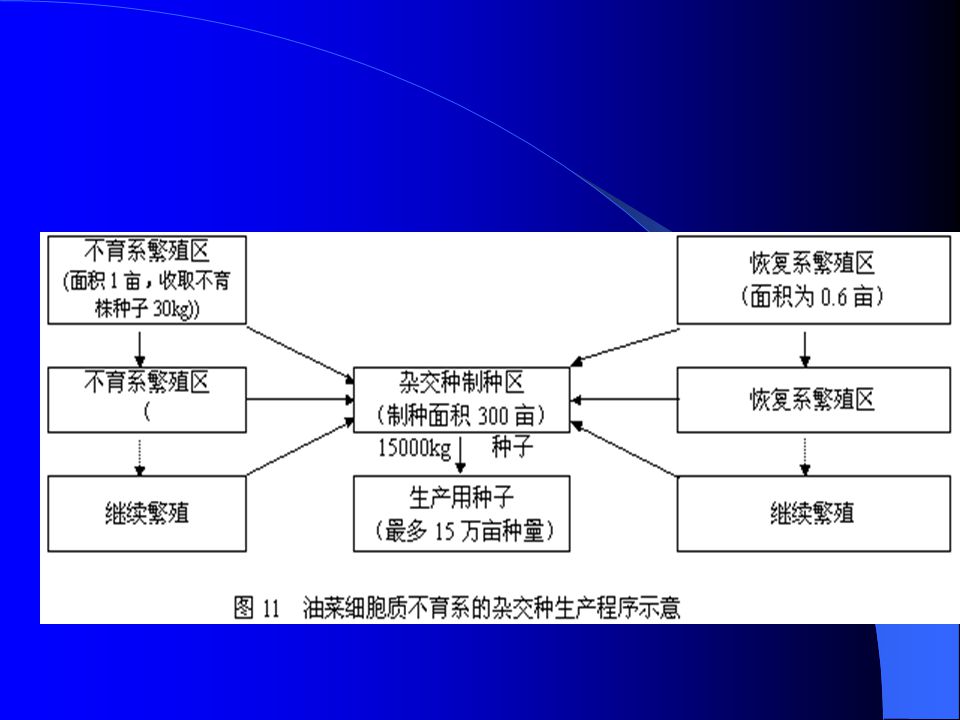
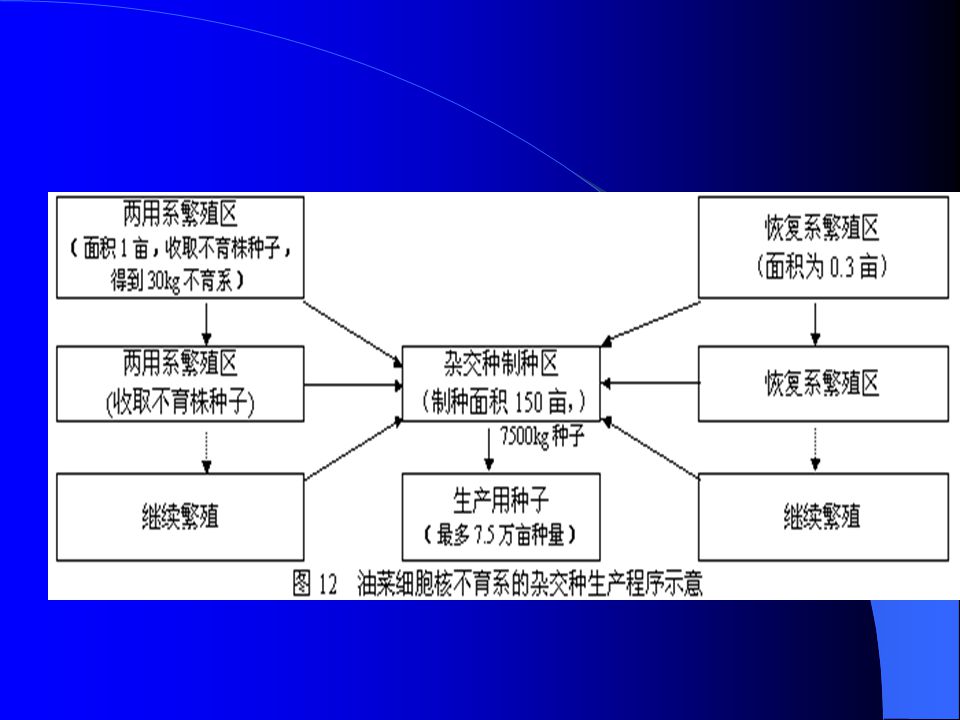
三、油菜杂交种选育 不育材料的来源及其遗传 1972 年英国 Thompson 和日本 Shiga 发现了细胞质与细胞核互作型 雄性不育材料,后合并称为 nap cms ,该不育材料在低温情况下不 育,温度高于 20 0 C 时,出现大量花粉,利用价值不大。同年,中 国傅庭栋在品种资源 Polima 中发现了 19 株天然不育株,分散给各 单位利用和研究,后该不育系统被命名为 Polima cms ,属于细胞 质与细胞核互作型雄性不育材料,被普遍认为是最有价值的雄性 不育基因资源。 1976 年,李殿荣在种间杂交组合 S74-3× (丰收 4 号 +7207 )中发现雄性不育系,经过回交、选择,育成了陕 2A cms 不育系和保持系,后育成了世界上第一个大规模应用的杂交种 “ 秦 油 2 号 ” ,标志着油菜杂种优势利用的突破性进展。
中发现雄性不育系,经过回交、选择,育成了陕 2A cms 不育系和保持系,后育成了世界上第一个大规模应用的杂交种 秦 油 2 号 ,标志着油菜杂种优势利用的突破性进展。")
22
油菜与播娘蒿融合

23
质核互作型不育系选育

25
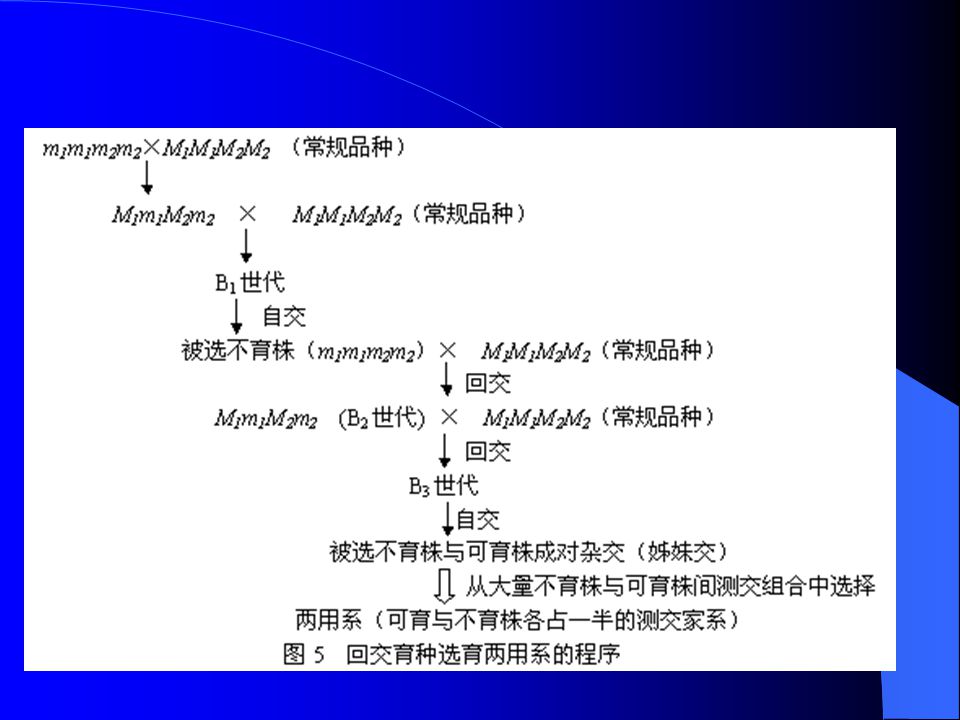
细胞核不育系选育

28
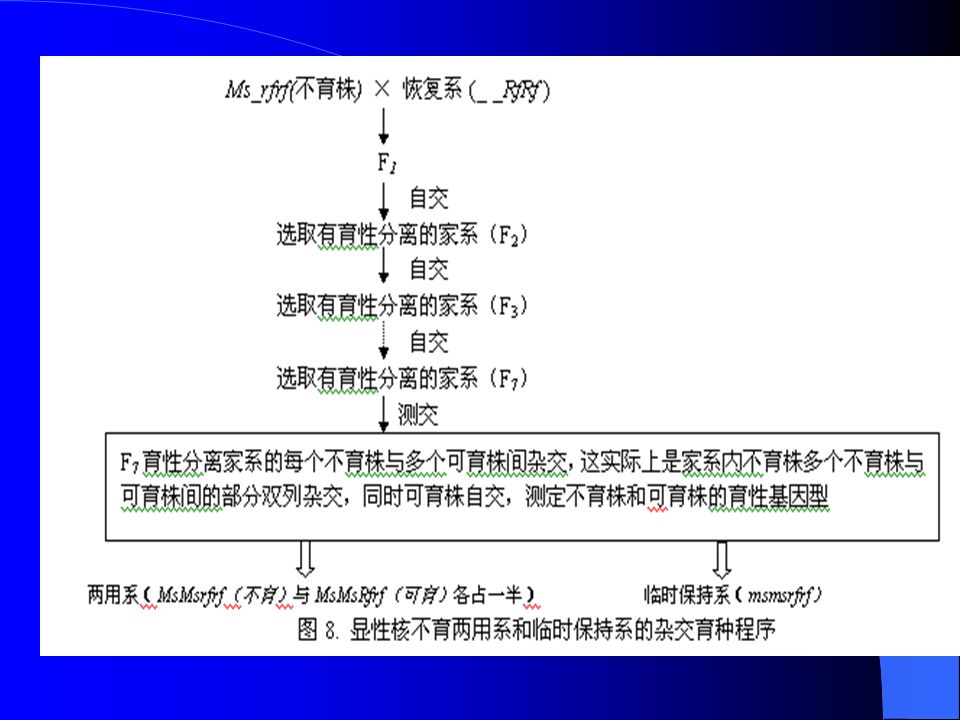
显性核不育系的选育

31
细胞核 + 细胞质不育系的选育

34
自交不亲和系的选育

35
转基因基因不育系的选育

36
化学杀雄杂交种 化学杀雄杂交种是用杀雄剂将母本的雄蕊杀死, 作为不育系,与常规品种杂交配制强优势组合 的方式生产的。最早有官春云等提出并研制了 杀雄剂 1 号,在单核花粉期喷施,造成花粉死 亡,雄性不育,制种时能够获得 50% 到 80% 的 杂种率。 化学杀雄利用杂种优势的方法,具有简便,易 于配制杂交组合,育种进程快的优点,但是杂 交种的纯度不易保证。

37
生态型不育系的选育

38
隐性上位性核不育系选育

39
四、杂交种制种技术 设置好隔离区,保证制种条件 调节开花期,保证父母本花期一致 保证亲本纯度 选择适当的行比 做好除杂、去劣工作 拔除野生十字花科植物、清理制种区 做好栽培管理 开花后砍除父本 人工辅助授粉

Podobne prezentacje







>")














![Typowe wartości dyn w różnych typach gwiazd: GWIAZDA [g/cm 3 ] dyn gwiazda neutronowa 10 15 0.06 ms Biały karzeł10 6 2 s Słońce1,4126 min Czerwony.](/11/3016618/big_thumb.jpg "Typowe wartości dyn w różnych typach gwiazd: GWIAZDA [g/cm 3 ] dyn gwiazda neutronowa 10 15 0.06 ms Biały karzeł10 6 2 s Słońce1,4126 min Czerwony.>")

