Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
Regulacja kwitnienia

2
Rośliny kwiatowe przechodzą fazę wzrostu wegetatywnego (wytwarzanie pędów i liści) i fazę kwitnienia, w trakcie której wytwarzają organy służące do rozmnażania płciowego U roślin jednorocznych faza wegetatywna zaczyna się w momencie kiełkowania nasion. Następująca po niej faza kwitnienia kończy się starzeniem się i śmiercią rośliny. U roślin dwuletnich, faza wegetatywna trwa przez pierwszy rok, w drugim roku następuje kwitnienie, które kończy się śmiercią rośliny. U roślin wieloletnich, kwitnienie następuje co rok, przez wiele lat. Wzrost wegetatywny pędu następuje w merystemie wierzchołkowym. Jest to masa niezróżnicowanych komórek na szczycie pędu. Podziały mitotyczne tych komórek wytwarzają komórki, które różnicują w części pędu, liście, wtórne merystemy (zwane też pączkami bocznymi) – dają początek rozgałęzieniom pędu.
 – dają początek rozgałęzieniom pędu.")
3
Kwitnienie jest regulowane przez wiele czynników
Kwitnienie wymaga przekształcenia merystemu wierzchołkowego w merystem kwiatowy Zależy to od: Czynników wewnętrznych Czynników zewnętrznych

4
Regulacja przez temperaturę
Wiele roślin jednorocznych (np. pszenica ozima) i dwuletnich ma opóźniony czas kwitnienia, jeśli nie przejdzie w trakcie zimy okresu zimowego przechłodzenia. Zmiany powodowane przez ten okres zimowego przechłodzenia noszą nazwę wernalizacji. U wielu drzewiastych roślin kwiatowych rosnących w klimacie umiarkowanym (jabłonie, bzy), kwitnienie wymaga uprzedniej ekspozycji na niską temperaturę. Drzewa te nie kwitną w klimacie ciepłym, w którym nie ma wyraźnych zim. Stan uśpienia pąków jest zlokalizowany.
 i dwuletnich ma opóźniony czas kwitnienia, jeśli nie przejdzie w trakcie zimy okresu zimowego przechłodzenia. Zmiany powodowane przez ten okres zimowego przechłodzenia noszą nazwę wernalizacji. U wielu drzewiastych roślin kwiatowych rosnących w klimacie umiarkowanym (jabłonie, bzy), kwitnienie wymaga uprzedniej ekspozycji na niską temperaturę. Drzewa te nie kwitną w klimacie ciepłym, w którym nie ma wyraźnych zim. Stan uśpienia pąków jest zlokalizowany.")
5
Regulacja przez fotoperiod (stosunek długości dnia do nocy)
Fotoperiod jest wykrywany w liściach (np. roślina X potrzebuje dnia o długości co najmniej 8,5 godziny, by móc zakwitnąć). Jednak wystarczy, by tylko jeden liść był eksponowany na właściwy fotoperiod, aby kwiaty pojawiały się na całej roślinie. Liście produkują sygnał chemiczny - florigen, który jest transportowany do merystemów wierzchołkowych. Chemiczna natura florigenu nie jest wyjaśniona, jednym z jego składników może być białko kodowane przez gen FT (Flowering locus T). Florigen może się przemieszczać poprzez system naczyniowy
. Jednak wystarczy, by tylko jeden liść był eksponowany na właściwy fotoperiod, aby kwiaty pojawiały się na całej roślinie. Liście produkują sygnał chemiczny - florigen, który jest transportowany do merystemów wierzchołkowych. Chemiczna natura florigenu nie jest wyjaśniona, jednym z jego składników może być białko kodowane przez gen FT (Flowering locus T). Florigen może się przemieszczać poprzez system naczyniowy.")
6
Budowa kwiatu Merystem kwiatowy różnicuje w cztery koncentryczne okółki (kręgi) komórek, które tworzą następnie cztery części kwiatu. Komórki w okółku 1 rozwijają się w działki kielicha, tworzące najniższy poziom. Łącznie działki tworzą tzw. kielich. Okółek 2 daje początek umieszczonym nad kielichem płatkom, tworzącym razem koronę kwiatu. Korona kwiatu jest jego najbardziej barwną częścią. Okółek 3 rozwija się w pręciki, męskie organy płciowe. Okółek 4 (najbardziej wewnętrzny) tworzy słupki, narządy płciowe żeńskie. Często zlewają się w pojedynczą strukturę. 1 2 3 4
 tworzy słupki, narządy płciowe żeńskie. Często zlewają się w pojedynczą strukturę")
7
Model ABC rozwoju kwiatu
Wyniki analizy genetycznej mutantów Arabidopsis i Petunii, sugerowały, że istnieje grupa genów kodujących czynniki transkrypcyjne (główne włączniki) niezbędne do włączania genów warunkujących rozwój działek kielicha, płatków korony, pręcików i słupków. Te główne włączniki należą do trzech klas: A, B i C. Komórki, w których wyrażane są tylko geny klasy A tworzą działki kielicha. Komórki, w których wyrażane są zarówno geny klasy A jak i klasy B, tworzą płatki korony. Komórki, w których wyrażane są zarówno geny klasy B jak i klasy C, tworzą pręciki. Komórki, w których wyrażane są tylko geny klasy C tworzą słupki. Geny ABC: Grupa A: Apetala1 (AP1) Apetala2 (AP2) Grupa B: Apetala3 (AP3) Pistilata (PI) Grupa C: Agamous (AG)
 niezbędne do włączania genów warunkujących rozwój działek kielicha, płatków korony, pręcików i słupków. Te główne włączniki należą do trzech klas: A, B i C. Komórki, w których wyrażane są tylko geny klasy A tworzą działki kielicha. Komórki, w których wyrażane są zarówno geny klasy A jak i klasy B, tworzą płatki korony. Komórki, w których wyrażane są zarówno geny klasy B jak i klasy C, tworzą pręciki. Komórki, w których wyrażane są tylko geny klasy C tworzą słupki. Geny ABC: Grupa A: Apetala1 (AP1) Apetala2 (AP2) Grupa B: Apetala3 (AP3) Pistilata (PI) Grupa C: Agamous (AG)")
8
Efekty mutacji w genach ABC

9
Relacje między genami ABC
Geny klasy A i C są w stosunku do siebie represorami. W nieobecności A, C są aktywne w całym kwiecie. W nieobecności C, A są aktywne w całym kwiecie. Efekty mutacji w AP1

10
Geny klasy E (SEP) uzupełniają geny modelu ABC
Geny SEP (SEPALLATA), obok genów ABC, są niezbędne do prawidłowego określania tożsamości organów kwiatowych
, obok genów ABC, są niezbędne do prawidłowego określania tożsamości organów kwiatowych.")
11
Większość genów ABCE koduje czynniki transkrypcyjne z domeną MADS
Domena MADS występuje na N-końcu białka i warunkuje: wiązanie do DNA, zdolność do dimeryzacji i lokalizację jądrową. (tylko białko AP2 nie należy do rodziny białek MADS) Arabidopsis zawiera ponad 100 genów kodujących różne białka z domeną MADS. Domena K MADS Domena C
 Arabidopsis zawiera ponad 100 genów kodujących różne białka z domeną MADS. Domena K. MADS. Domena C.")
12
Niezależna ewolucja genetycznych narzędzi kontroli rozwoju u zwierząt i roślin
Meyerowitz, EM. Science (2002)
")
13
Centralna rola genu LFY (LEAFY)
Ortologi LFY występują u wszystkich gatunków roślin (także nie kwiatowych). Aktywność LFY jest konieczna i wystarczająca do determinacji merystemu kwiatowego. Niezależnie od determinacji typu merystemu, LFY pełni dwie kluczowe funkcje w rozwoju kwiatu: Jest głównym integratorem sygnałów prowadzących do indukcji kwitnienia. Jest głównym aktywatorem genów ABCE.
. Aktywność LFY jest konieczna i wystarczająca do determinacji merystemu kwiatowego. Niezależnie od determinacji typu merystemu, LFY pełni dwie kluczowe funkcje w rozwoju kwiatu: Jest głównym integratorem sygnałów prowadzących do indukcji kwitnienia. Jest głównym aktywatorem genów ABCE.")
14
Cztery fazy rozwoju kwiatu
W odpowiedzi na sygnały ze środowiska i sygnały wewnętrzne roślina przestawia się z wzrostu wegetatywnego na wzrost reprodukcyjny – ten proces kontrolują geny regulujące czas kwitnienia (flowering time genes). Sygnały z różnych ścieżek wpływających na czas kwitnienia są integrowane, co prowadzi do aktywacji niewielkiej grupy genów tożsamości merystemu (meristem identity genes), które warunkują powstanie kwiatu (Geny TFL1, LFY, AP1). Geny tożsamości merystemu aktywują geny tożsamości organów kwiatowych (Geny ABCE). Geny tożsamości organów aktywują zależne od nich geny „budujące organy”, które determinują różne typy komórek, z których składają się poszczególne organy kwiatu.
. Sygnały z różnych ścieżek wpływających na czas kwitnienia są integrowane, co prowadzi do aktywacji niewielkiej grupy genów tożsamości merystemu (meristem identity genes), które warunkują powstanie kwiatu (Geny TFL1, LFY, AP1). Geny tożsamości merystemu aktywują geny tożsamości organów kwiatowych (Geny ABCE). Geny tożsamości organów aktywują zależne od nich geny „budujące organy , które determinują różne typy komórek, z których składają się poszczególne organy kwiatu.")
15
Genetyczna kontrola czasu kwitnienia
Geny kontrolujące czas kwitnienia działają w czterech ścieżkach indukcji: Zależnej od fotoperiodu (rytm dobowy, długi dzień); B Zależnej od giberelin (GA); C Autonomicznej; D Wernalizacyjnej
; B. Zależnej od giberelin (GA); C. Autonomicznej; D. Wernalizacyjnej.")
16
U Arabidopsis liczba liści w rozecie jest miarą czasu kwitnienia
In Arabidopsis vegetative phase of development - rosette, generative phase -stem with flowers and seeds FLC is a repressor of transition to flowering. FLC is up-regulated in late flowering mutants - efficiently repressing transition to flowering

17
Analiza czasu kwitnienia w Arabidopsis, efekt mutacji w genach SWI3
% days leaf No C /27 D /34 Wt /54 days leaf No C /9-10 D /11 Wt /11 C-3-4 D-6-7 Wt-8 C-3-4 D-6-7 Wt-8 Sarnowski et al.,Plant Cell 2005 atswi3c: early flowering in SD, slightly early flowering in LD atswi3d: early flowering in SD

18
Kontrola czasu kwitnienia – fotoperiod (long day pathway)
Wiele genów indukujących kwitnienie w długim dniu koduje białka zaangażowane w percepcję światła (PHYTOCHROME A, CRYPTOCHROM2) lub składniki regulujące zegar okołodobowy (GIGANTEA, ELF3). Geny te ostatecznie aktywują gen CO (CONSTANS) (mutany co są późnokwitnące, nadekspresja CO powoduje wczesne kwitnienie). CO koduje białko jądrowe zawierające dwie domeny palców cynkowych. AP1 LFY
 lub składniki regulujące zegar okołodobowy (GIGANTEA, ELF3). Geny te ostatecznie aktywują gen CO (CONSTANS) (mutany co są późnokwitnące, nadekspresja CO powoduje wczesne kwitnienie). CO koduje białko jądrowe zawierające dwie domeny palców cynkowych. AP1. LFY.")
19
Kontrola czasu kwitnienia – gibereliny
Mutanty z defektem w biosyntezie giberelin (np. ga1) są bardzo późno kwitnące w krótkim dniu, ale nie w długim, co wskazuje, że szlak GA ma kluczowe znaczenie w indukcji kwitnienia w sytuacji braku sygnału indukującego długiego dnia (fotoperiodycznego) AP1 LFY
 są bardzo późno kwitnące w krótkim dniu, ale nie w długim, co wskazuje, że szlak GA ma kluczowe znaczenie w indukcji kwitnienia w sytuacji braku sygnału indukującego długiego dnia (fotoperiodycznego) AP1. LFY.")
20
Kontrola czasu kwitnienia – szlak autonomiczny i szlak wernalizacyjny
Geny szlaku autonomicznego kontrolują kwitnienie niezależne od długiego dnia (Arabidopsis jest normalnie rośliną kwitnącą w długim dniu, ale po dłuższym okresie zakwita także w krótkim dniu). Kwitniecie w krótkim dniu jest indukowane przez geny szlaku autonomicznego. Geny szlaku wernalizacyjnego są związane z indukcją kwitnienia związana z przejściem zimowego przechłodzenia. Geny FLC, SOC1, FT i LFY pełnią funkcję integratorów sygnałów z różnych szlaków kontroli czasu kwitnienia. AP1 LFY
. Kwitniecie w krótkim dniu jest indukowane przez geny szlaku autonomicznego. Geny szlaku wernalizacyjnego są związane z indukcją kwitnienia związana z przejściem zimowego przechłodzenia. Geny FLC, SOC1, FT i LFY pełnią funkcję integratorów sygnałów z różnych szlaków kontroli czasu kwitnienia. AP1. LFY.")
21
Regulacja czasu kwitnienia
Przykład badań wybór właściwego czasu kwitnienia jest kluczowy dla sukcesu reprodukcyjnego roślin ewolucja doprowadziła do powstania wielu ścieżek regulujących czas kwitnienia

22
Kontrola czasu kwitnienia
Czynniki wewnętrzne Czynniki zewnętrzne Ścieżka zależna od GA Wernalizacja Ścieżka Autonomiczna Fotoperiod temperatura

23
FLC (Flowering Locus C) jest centralnym regulatorem kwitnienia
In Arabidopsis vegetative phase of development - rosette, generative phase -stem with flowers and seeds FLC is a repressor of transition to flowering. FLC is up-regulated in late flowering mutants - efficiently repressing transition to flowering

24
Struktura genu FLC ATG TAG gen FLC FLC is MADS box type transcription factor with 7 exons. showing homology to AGAMOUS (MADS box) in terms of sequence and in terms of it regulation. FLC is a well studied gen. Offers possibility to add new layers of regulation to already discovered.
 in terms of sequence and in terms of it regulation. FLC is a well studied gen. Offers possibility to add new layers of regulation to already discovered.")
25
mRNA genu FLC jest podwyższony w późno kwitnących ekotypach Arabidopsis
FLC is highly expressed in late flowering Arabidopsis mutants and in late Arabidopsis Ecotypes. Nature uses FLC to repress flowering.

26
FLC podlega supresji przez Ścieżkę Autonomiczną
Autonomiczna FCA, FY FLC FLC is suppressed by Autonomous pathway mutants.

27
FCA FY RBD WW Q białko wiążące RNA Związane z obróbką 3` końca
transkryptów FY rejon WD silnie konserwowany u wszystkich eukariontów WD PPLP

28
model działania kompleksu FCA & FY
PPLP FY pA/cleavage complex WD40 sygnały poliadenylacji RNP WW FCA transcripty -FLC ? -FCA SWI3 It terns out that studding FLC regulation we can get closer to understand general biological questions -like mRNA 3` end processing.

29
czy FLC jest regulowany przez RNAi
? RNAi FLC

30
RNAi (RNA interference)
cechą charakterystyczną wszystkich odmian RNAi są małe 21-24nt RNA RNAi powstało najprawdopodobniej jako mechanizm obrony przeciw wirusom, występuje u wszystkich badanych eukariontów RNAi został wykorzystany przez komórki do utrzymywania heterochromatyny na cetromerach i obszarach zawierających wielokrotne powtórzenia odmianą RNAi używaną przez wszystkie znane eukarionty wielokomórkowe jest RNAi z udziałem miRNA sRNA first discovered in A. thaliana

31
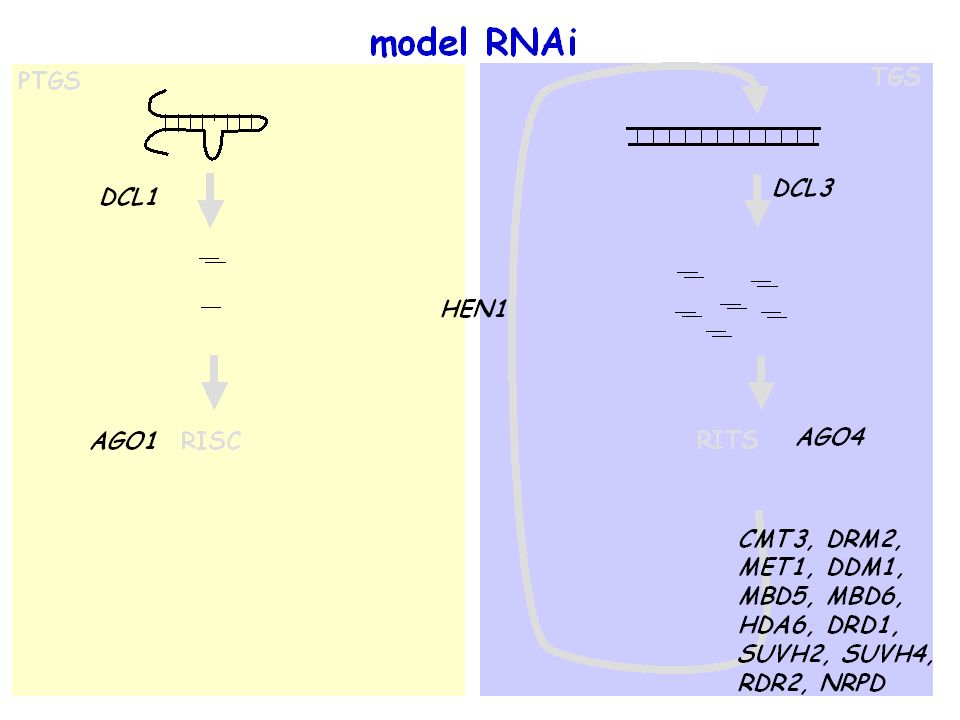
model RNAi miRNA siRNA PTGS (Post Transcriptional Gene Silencing) TGS
prekursor dsRNA 21-24 nt małe RNA miRNA siRNA RISC (RNA induced Silencing Complex) RITS (RNA induced Transcriptional Silencing) tworzenie heterochromatyny: metylacja DNA, metylacja histonu H3 K9 cięcie mRNA lub inhibicja translacji
 RITS. (RNA induced Transcriptional Silencing) tworzenie heterochromatyny: metylacja DNA, metylacja histonu H3 K9. cięcie mRNA lub inhibicja translacji.")
32
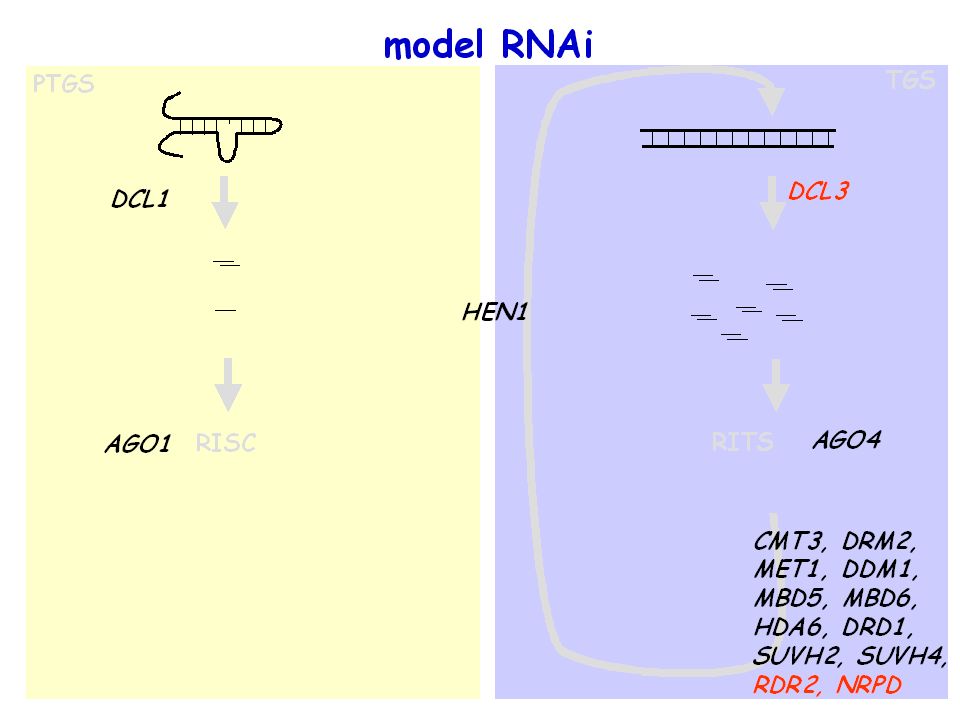
model RNAi PTGS (Post Transcriptional Gene Silencing) TGS
prekursor dsRNA 21-24 nt małe RNA RISC (RNA induced Silencing Complex) RITS (RNA induced Transcriptional Silencing) regulacja ekspresji genów (30% ludzkich genów) Integralność centromerów, kontrolowanie transpozonów, regulacja ekspresji genów?
 RITS. (RNA induced Transcriptional Silencing) regulacja ekspresji genów. (30% ludzkich genów) Integralność centromerów, kontrolowanie transpozonów, regulacja ekspresji genów")
34
czy FLC jest regulowany przez RNAi
? RNAi FLC

35
nowe małe RNA o sekwencji homologicznej do rejonu 3`FLC
ATG TAG użyta sonda liście rozety łuszczynki liście pędu kwiaty siewki pęd Looked for small RNA with homology to FLC genomic sequence. Found sRNA with homology to region precisely outside FLC 3`UTR. Small RNA expressed in siliques. sonda do rejonu 3`FLC 30nt 20nt EtBR

37
poziom małych RNA z 3`FLC jest obniżony w mutantach ze ścieżki polIV
Ler Col-0 dcl 2 dcl 3 sgs2/rdr6 nrpd1a-3 nrpd1a-2 rdr2 ago4-1 30nt 20nt

38
model RNAi regulacja FLC przez RNAi– pierwszym przykładem regulacji
PTGS TGS RISC RITS Integralność centromerów, kontrolowanie transpozonów, regulacja ekspresji genów? regulacja FLC przez RNAi– pierwszym przykładem regulacji jednokopijnego genu nie związanego z transpozonami przez siRNA

39
3`koniec FLC jest związany z transkrypcją w orientacji sens i antysens
small RNA region ATG TAG orientacja sens antysens To study the transcription at the 3` end of FLC we used whole genome array data kindly provided by..(joł).. Ecker before publication. The data was obtained by hybridising polyA RNA from different Arabidopsis tissue to 25 oligonucleotides spotted on an array covering the whole Arabidopsis genome. Massive transcription in sense orientation. single antisense pick from inflorescence corresponding to my small RNA. Yamada et al. Science 2003
.. Ecker before publication. The data was obtained by hybridising polyA RNA from different Arabidopsis tissue to 25 oligonucleotides spotted on an array covering the whole Arabidopsis genome. Massive transcription in sense orientation. single antisense pick from inflorescence corresponding to my small RNA. Yamada et al. Science")
40
mutanty T-DNA w rejonie 3`FLC
flc_utr5 flc_utr4 flc_utr3 rejon małych RNA ATG TAG

41
Insercja w rejon 3`FLC zaburza negatywną regulację FLC
ilość liści w warunkach długiego dnia poziom mRNA FLC 3 independent Northern showed a average increase of 20% in FLC mRNA level in utr4 mutant. poziom mRNA B- tubuliny Col-0 utr 3 utr 4 utr 5

42
FLC podlega supresji przez mutanty ścieżki RNAi
Col dcl2-1 dcl3-1 rdr2-1 sde4-2 sde4-3 ilość liści w krótkim dniu All mutants loosing FLC 3` sRNA show increased FLC mRNA levels but not for example dcl2 or ago4 (data not shown).
.")
43
Struktura chromatyny

44
Struktura chromatyny euchromatyna heterochromatyna H3 K9 diME

45
metoda ChIP (Chromatin Immuno Precipitation)
przeciwciała specyficzne do H3 di-meK9 Experimental method for analyzing the acetylation state of histones in chromatin associated with a specific region of the genome Nucleosomes are lightly cross-linked to DNA in vivo using a cell-premeable, reversible, chemical cross-linking agent. Chromatin is then isolated, sheared to an average length of three nucleosomes, and subjected to immunoprecipitation with an antibody specific for a particular acetylated N-terminal histone sequence. The DNA in the immunoprecipitated chromatin fragment is released by reversing the cross-link and then is quantitated using a sensitive PCR method. Basically at the end of this you can get an idea which genomic regions were associated with histones (as a nucleosome) that were modified. amplifikacja PCR wzbogaconych fragmentów
 that were modified. amplifikacja PCR. wzbogaconych fragmentów.")
46
metoda ChIP ???? Jenuwein T

47
W łuszczynkach rejon 3`FLC jest związany ze znacznikiem H3 K9 di-methyl
NoAB ChIP ŁUSZCZYNKI ŁUSZCZYNKI SIEWKI SIEWKI Ta3 rejon 3`FLC

48
znacznik H3 K9 di-methyl jest ograniczony do rejonu 3`FLC
ATG TAG Wzbogacenie w stosunku do kontroli Wzbogacenie zanaczonych reionów w H3 di-me K9; ChIP nad Input

49
regulacja FLC RNAi FLC

50
regulacja FLC FRI Ścieżka Autonomiczna FCA, FY Wernalizacja RNAi FLC

51
Świeżewski, S. , Crevillen, P. , Liu, F. , Ecker, J. R
Świeżewski, S., Crevillen, P., Liu, F., Ecker, J.R., Jerzmanowski, A. and Dean, C. (2007) Small RNA-mediated chromatin silencing directed to the 3’ region of the Arabidopsis gene encoding the developmental regulator, FLC. Proc. Natl. Acad. Sci. USA 104,
 Small RNA-mediated chromatin silencing directed to the 3’ region of the Arabidopsis gene encoding the developmental regulator, FLC. Proc. Natl. Acad. Sci. USA 104,")
Podobne prezentacje