Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
Replikacja u Eukaryota
Proces replikacji u Eukaryota przebiega bardzo podobnie do replikacji w komórkach bakteryjnych. Zasadniczą różnicą jest istnienie wielu ori. U drożdży podobnie jak u bakterii ori (nazywane tu ARS od ang. autonomously replicating sequence) zawiera sekwencje bogate w A i T: [A/T]TTTAT[A/G]TTT[A/T]
 zawiera sekwencje bogate w A i T: [A/T]TTTAT[A/G]TTT[A/T]")
2
WAWTTDDWWWDHWGWHMAWTT
U człowieka jest ok ori. Znajdują się co 30 – 300 kpz. NIE SĄ DOBRZE ZDEFINIOWANE Komputerowa analiza wielu potencjalnych ori pozwoliła na zaproponowanie takiej sekwencji konsensusowej: WAWTTDDWWWDHWGWHMAWTT gdzie: W = A lub T; D = A lub G; H = A lub T lub C; M = A lub C.

3
Jak można badać replikację u Eukaryota?
Replikacja w układzie bezkomórkowym – replikacja wirusów DNA np. wirusa SV40 (małpi wirus) lub wirusa brodawczaka bydlęcego (papilloma). Wyłączanie aktywności niektórych białek przez przeciwciała Model drożdżowy – badanie mutantów
 lub wirusa brodawczaka bydlęcego (papilloma). Wyłączanie aktywności niektórych białek przez przeciwciała. Model drożdżowy – badanie mutantów.")
4
Inicjacja replikacji:
Do ori przyłącza się kompleks ORC złożony z 6 białek (o m. cz. ~100, 82, 66, 50, 50, 28 kDa) ORC – (ang. origin of replication complex) ORC pełni tę samą funkcję co DnaA u bakterii Następnie helikaza rozplata helisę, a polimeraza a syntetyzuje startery na nici wiodącej i opóźnionej. Funkcję helikazy pełni kompleks białek MCM 2-7 (ang. minichromosome maintenance) = DNA replication licencing factors (tzw. czynniki upoważniające).
 ORC – (ang. origin of replication complex) ORC pełni tę samą funkcję co DnaA u bakterii. Następnie helikaza rozplata helisę, a polimeraza a syntetyzuje startery na nici wiodącej i opóźnionej. Funkcję helikazy pełni kompleks białek MCM 2-7 (ang. minichromosome maintenance) = DNA replication licencing factors (tzw. czynniki upoważniające).")
5
POLIMERAZA a Składa się z czterech podjednostek:
Podjednostka 180 kDa – aktywność polimerazy DNA Podjednostki 55 kDa i 48 kDa – tworzą razem prymazę (polimeraza RNA). Podjednostka 68 kDa spaja kompleks a może mieć też znaczenie regulatorowe (ulega fosforylacji). Syntetyzuje startery złożone z 10 nukleotydów RNA (prymaza) i nukleotydów DNA (polimeraza) NIE MA AKTYWNOŚCI EGZONUKLEAZY
. Podjednostka 68 kDa spaja kompleks a może mieć też znaczenie regulatorowe (ulega fosforylacji). Syntetyzuje startery złożone z 10 nukleotydów RNA (prymaza) i nukleotydów DNA (polimeraza) NIE MA AKTYWNOŚCI EGZONUKLEAZY.")
6
POLIMERAZY d oraz e Głównymi polimerazami replikacyjnymi Eukaryota są polimerazy delta i epsilon (d i e). Polimerazy d i e są złożone z 4 podjednostek o różnych masach cząsteczkowych: - (p125, p66, p50, p12) e - (p256, p80, p23, p22) Największe podjednostki obu polimeraz zawierają zarówno aktywność polimerazy jak i 3’-5’ egzonukleazy. Polimeraza d odpowiada za replikację nici opóźnionej Polimeraza e odpowiada za replikację nici wiodącej Uwaga! W podręczniku „Zarys biochemii” jest informacja nieaktualna!
 e - (p256, p80, p23, p22) Największe podjednostki obu polimeraz zawierają zarówno aktywność polimerazy jak i 3’-5’ egzonukleazy. Polimeraza d odpowiada za replikację nici opóźnionej. Polimeraza e odpowiada za replikację nici wiodącej. Uwaga! W podręczniku „Zarys biochemii jest informacja nieaktualna!")
7
Analogicznie do Prokaryota – polimeraza replikacyjna musi oddziaływać ze strukturą, która będzie zwiększać jej kontakt z DNA, będzie zwiększać jej procesywność. Tą strukturą u Eukaryota jest PCNA (ang. proliferating cell nuclear antigen), antygen jądrowy proliferujących komórek. Trimer PCNA 3 x 29 kDa Oddziaływanie PCNA z DNA i kompleksem polimerazy reguluje RFC (replikacyjny czynnik C)
")
8
U EUKARYOTA NIE MA WIĘC ODPOWIEDNIKA PROKARIOTYCZNEJ POLIMERAZY I
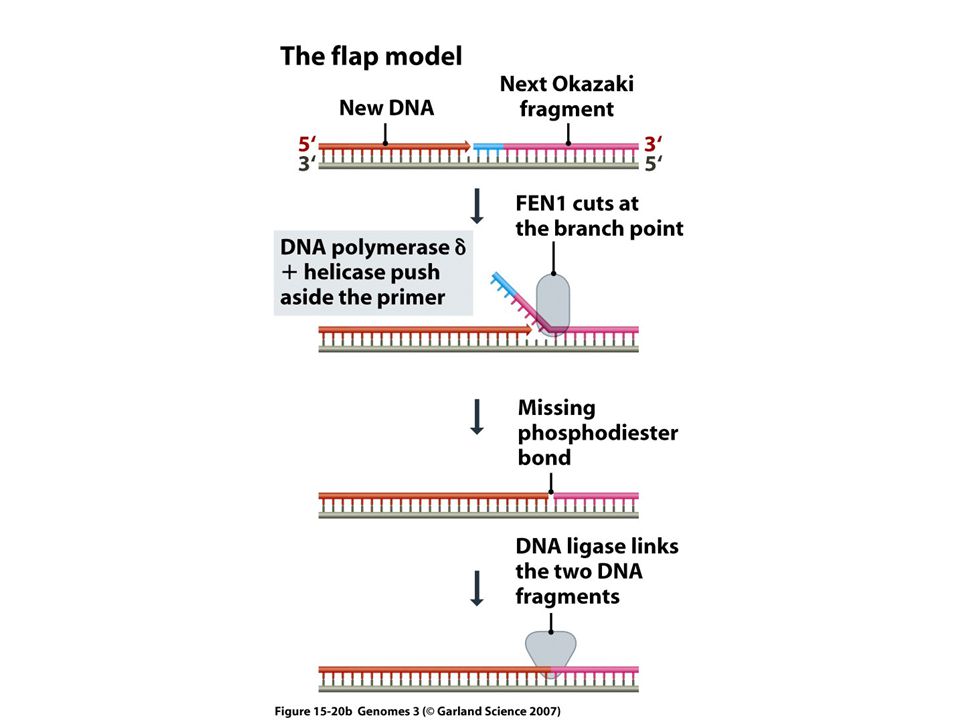
W trakcie syntezy nici opóźnionej: Synteza fragmentu Okazaki zostaje zatrzymana, gdy polimeraza d napotka starter poprzedniego fragmentu Okazaki. Wtedy następuje usunięcie tego startera przez działanie dwóch enzymów: Dna2 (DNA helikaza/endonukleaza 2) oraz endonukleazę Fen-1 (Flap endonuclease). Luka zostaje wypełniona przez polimerazę d. U EUKARYOTA NIE MA WIĘC ODPOWIEDNIKA PROKARIOTYCZNEJ POLIMERAZY I
 oraz endonukleazę Fen-1 (Flap endonuclease). Luka zostaje wypełniona przez polimerazę d. U EUKARYOTA NIE MA WIĘC ODPOWIEDNIKA PROKARIOTYCZNEJ POLIMERAZY I.")
10
ORC (ang. origin of replication complex);
FUNKCJA E. coli Człowiek Inicjacja replikacji DnaA ORC Synteza nici wiodącej i opóźnionej polimeraza DNA III polimerazy e i d Procesywność podjednostka b PCNA Kontrola oddziaływania b lub PCNA z DNA kompleks g RFC Prymaza DnaG polimeraza a Helikaza DnaB kompleks MCM2-7 ochrona ssDNA SSB RPA Topologia gyraza Topo I i Topo II ORC (ang. origin of replication complex); PCNA (ang. proliferating cell nuclear antigen); antygen jądrowy proliferujących komórek RFC (ang. replication factor C); replikacyjny czynnik C RPA (ang. replication protein A); replikacyjne białko A MCM (ang. minichromosome maintenance)
; PCNA (ang. proliferating cell nuclear antigen); antygen jądrowy proliferujących komórek. RFC (ang. replication factor C); replikacyjny czynnik C. RPA (ang. replication protein A); replikacyjne białko A. MCM (ang. minichromosome maintenance)")
11
Przykłady polimeraz DNA u ssaków

12
Polimerazy eta oraz iota należą do rodziny Y polimeraz DNA.
Uczestniczą w tzw. „translesion DNA synthesis (TLS)” ang. lesion = rana, uszkodzenie To są bardzo ciekawe polimerazy. Pracują na matrycy, ale nie dbają zbytnio o wierność replikacji. Dzięki temu mogą prowadzić replikację nawet wtedy, gdy matryca zawiera błędy (polimeraza d w tej sytuacji nie jest w stanie włączać nukleotydów do łańcucha DNA). Błędy wprowadzane przez polimerazy Y są później naprawiane przez różne systemy naprawy DNA.
 ang. lesion = rana, uszkodzenie. To są bardzo ciekawe polimerazy. Pracują na matrycy, ale nie dbają zbytnio o wierność replikacji. Dzięki temu mogą prowadzić replikację nawet wtedy, gdy matryca zawiera błędy (polimeraza d w tej sytuacji nie jest w stanie włączać nukleotydów do łańcucha DNA). Błędy wprowadzane przez polimerazy Y są później naprawiane przez różne systemy naprawy DNA.")
13
Polka współodkrywcą elementu mechanizmu naprawy DNA
PAP, 24-letnia Polka kierowała międzynarodowym projektem badawczym, w ramach którego dokonano bardzo ważnego odkrycia w dziedzinie biologii molekularnej - ustalono nieznany wcześniej element jednego z podstawowych mechanizmów naprawy uszkodzonego DNA. Science 16, December 2005: Vol no. 5755, pp – 1824 Ubiquitin-Binding Domains in Y-Family Polymerases Regulate Translesion Synthesis Marzena Bienko, Catherine M. Green, Nicola Crosetto, Fabian Rudolf, Grzegorz Zapart, Barry Coull, Patricia Kannouche, Gerhard Wider, Matthias Peter, Alan R. Lehmann, Kay Hofmann, Ivan Dikic

14
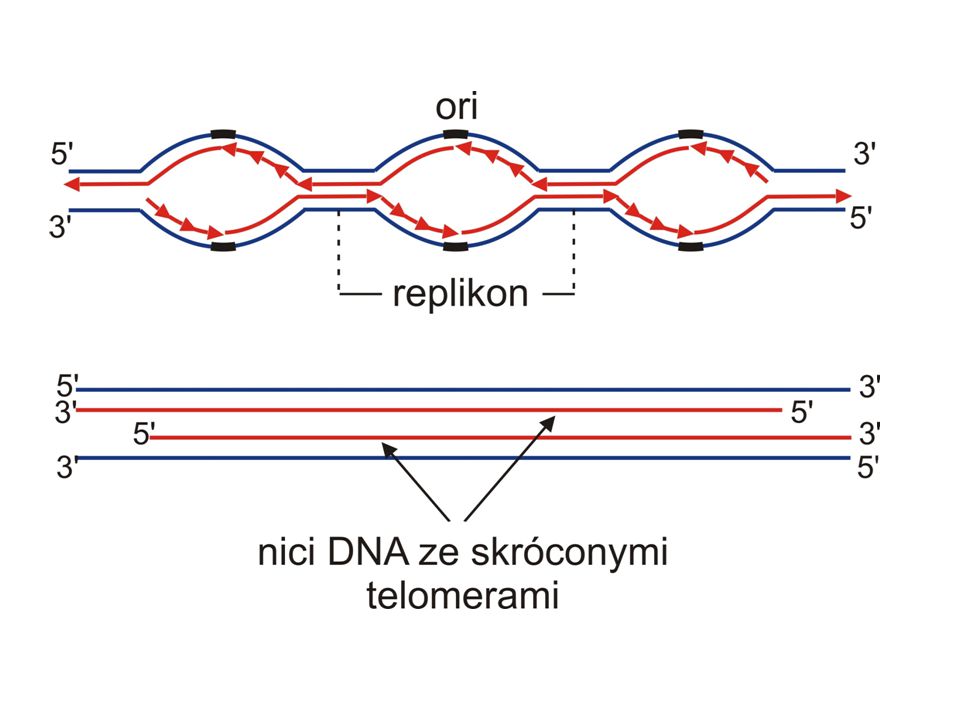
Jak może się skończyć synteza nici opóźnionej w liniowych chromosomach?

16
5’ koniec nowo syntetyzowanego DNA jest zawsze krótszy od macierzystego DNA
W nowej cząsteczce DNA fragment 3’ końca jest jednoniciowy (pojedyncza nić to 50 – 400 n) DWA PROBLEMY Jednoniciowy DNA jest bardzo podatny na degradację Przy każdym podziale komórkowym chromosomy ulegałyby (ulegają) skróceniu
 DWA PROBLEMY. Jednoniciowy DNA jest bardzo podatny na degradację. Przy każdym podziale komórkowym chromosomy ulegałyby (ulegają) skróceniu.")
17
w dziedzinie medycyny i fizjologii
NAGRODA NOBLA 2009 w dziedzinie medycyny i fizjologii Carol W. Greider Jack W. Szostak Elizabeth H. Blackburn for the discovery of how chromosomes are protected by telomeres and the enzyme telomerase

18
Telomery tworzą duże dwuniciowe pętle
TRF2 (ang. telomeric repeat binding factor 2) Obecnie wiadomo, że kompleks w skład, którego wchodzi TRF2 ma skomplikowaną strukturę
 Obecnie wiadomo, że kompleks w skład, którego wchodzi TRF2 ma skomplikowaną strukturę.")
19
ten kompleks nazywa się z ang. shelterin

20
Jak może się skończyć synteza nici opóźnionej w liniowych chromosomach?
Dzięki specyficznej budowie końców chromosomów – telomerów i enzymowi – telomerazie. DNA telomerów zawiera setki powtórzeń krótkich sekwencji bogatych w guanylan. Np. Tetrahymena (Orzęsek): GGGTTG Caenorhabditis (Nicień): GGCTTA Człowiek: GGGTTA 5’..... GGGTTAGGGTTAGGGTTAGGGTTAGGGTTA-3’ 3’......CCCAATCCCAAT-5’
: GGGTTG. Caenorhabditis (Nicień): GGCTTA. Człowiek: GGGTTA. 5’..... GGGTTAGGGTTAGGGTTAGGGTTAGGGTTA-3’ 3’......CCCAATCCCAAT-5’")
21
FISH fluorescent in situ hybridization
Fluorescencyjna hybrydyzacja in situ

22
5’..... GGGTTAGGGTTAGGGTTAGGGTTAGGGTTA-3’ 3’......CCCAATCCCAAT-5’
par zasad 400 zasad długość telomeru długość odcinka jednoniciowego Bioorganic and Medicinal Chemistry, 17, 5, 2009,

23
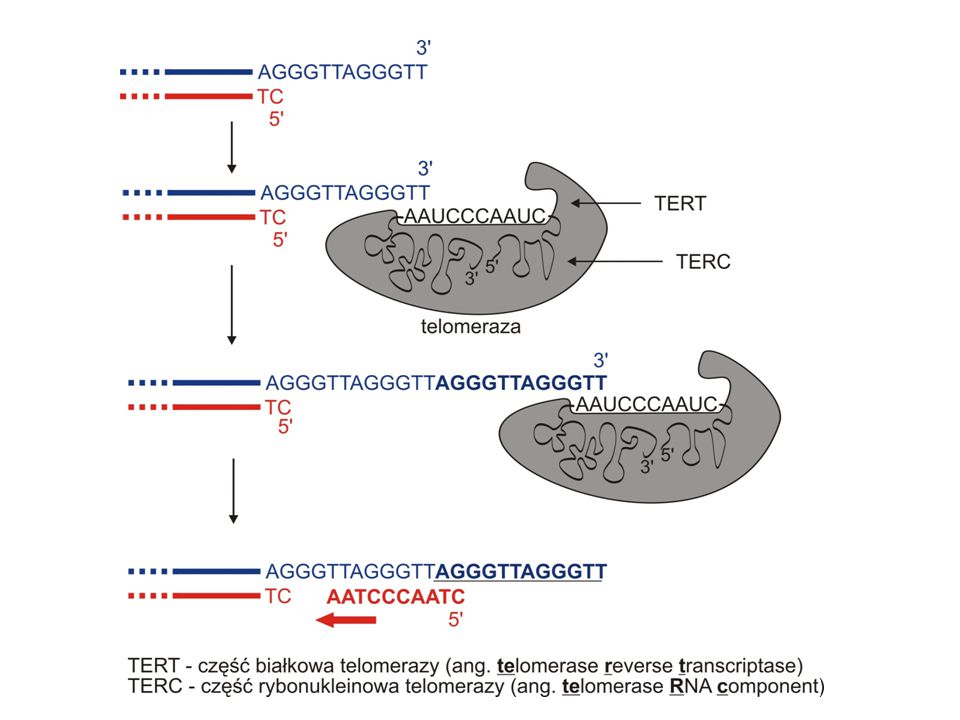
TELOMERAZA – rybonukleoproteina
Enzym o aktywności odwrotnej transkryptazy (ang. telomerase reverse transcriptase) U człowieka: TERT + odcinek RNA (TERC) + 8 białek pomocniczych
 U człowieka: TERT + odcinek RNA (TERC) + 8 białek pomocniczych.")
24
RNA telomerazy orzęska

26
przyłączenie telomerazy do telomeru
Etapy: przyłączenie telomerazy do telomeru wydłużenie końca 3' (nić macierzysta) przez telomerazę - synteza DNA na matrycy RNA wydłużenie nici opóźnionej przez polimerazę DNA - synteza DNA na matrycy DNA Telomeraza każdorazowo dodaje wiele powtórzeń charakterystycznej sekwencji telomerowej.
 przez telomerazę - synteza DNA na matrycy RNA. wydłużenie nici opóźnionej przez polimerazę DNA - synteza DNA na matrycy DNA. Telomeraza każdorazowo dodaje wiele powtórzeń charakterystycznej sekwencji telomerowej.")
27
a procesy starzenia się i nowotworzenia
Telomeraza i telomery a procesy starzenia się i nowotworzenia Hipoteza – skracanie telomerów warunkuje proces starzenia się? Czy długość telomerów determinuje długość życia? Porównując gatunki – NIE (mysz - długie telomery, człowiek – krótkie telomery) Czy ma wpływ na długość życia osobników w obrębie gatunku – NIE WIADOMO
 Czy ma wpływ na długość życia osobników w obrębie gatunku – NIE WIADOMO.")
28
W wielu tkankach wraz z wiekem osobniczym dochodzi do skracania telomerów. Im człowiek starszy, tym ma krótsze telomery. Co z tego wynika? NIC Równoległe zachodzenie 2 procesów nie determinuje zależności między nimi. Nie wiadomo, czy istnieje jakaś zależność, a jeśli tak, to który proces jest przyczyną, a który skutkiem. Np. starzenie i siwienie i zmarszczki. Czy można powiedzieć, że starzenie jest rezultatem siwienia i pojawiania się zmarszczek?

29
W hodowlach komórkowych istnieje zależność między aktywnością telomerazy a zdolnością komórek do proliferacji. Normalne komórki wykazują ograniczoną zdolność do podziałów. Po pewnej liczbie podziałów przestają proliferować – starzenie komórkowe.

30
Wiele tkanek dorosłego człowieka zachowuje aktywność telomerazy
Wiele tkanek dorosłego człowieka zachowuje aktywność telomerazy. Jakie to tkanki? Te, które muszą intensywnie proliferować: Komórki warstwy podstawnej naskórka Komórki nabłonka jelit Aktywowane limfocyty Komórki warstwy podstawnej endometrium (błony śluzowej macicy) (zależnie od fazy cyklu miesięcznego) Komórki mieszka włosowego
 (zależnie od fazy cyklu miesięcznego) Komórki mieszka włosowego.")
31
Telomeraza i nowotwory
Aktywność telomerazy nie stymuluje nowotworzenia jednak Krótkie telomery i brak aktywności telomerazy mogą hamować rozwój nowotworów. Podczas nowotworzenia, po serii podziałów komórkowych zmutowane komórki nie mogą dalej się dzielić, chyba, że zostanie przywrócona ekspresja telomerazy lub uruchomiony inny mechanizm wydłużania telomerów. Wiele – ale nie wszystkie! – nowotwory mają aktywność telomerazy.

32
USZKODZENIA I NAPRAWA DNA

34
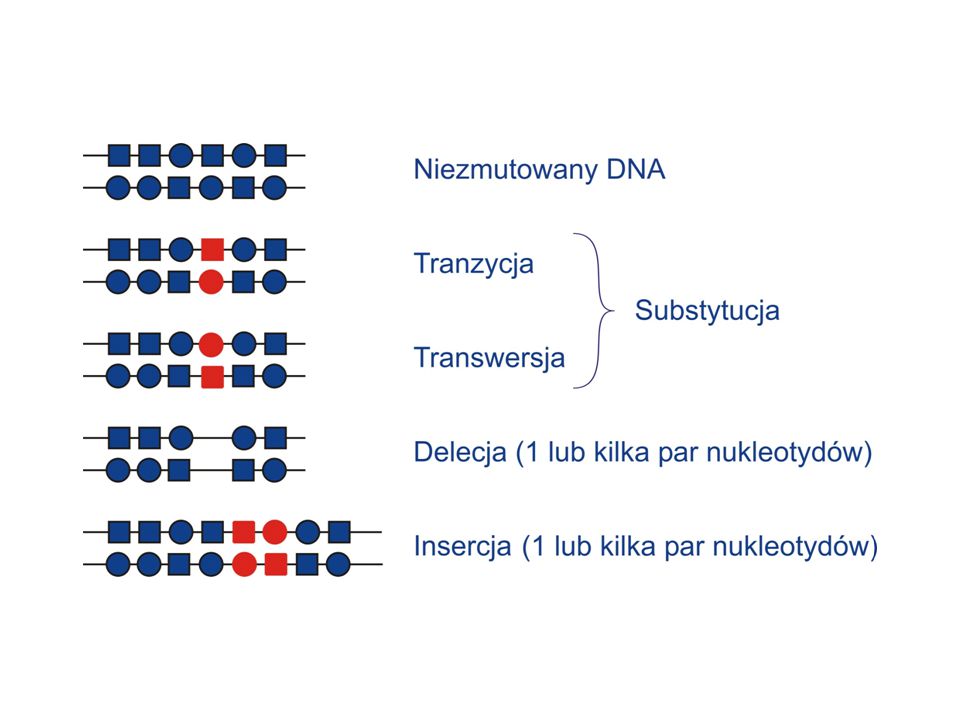
Część mutacji może powstawać w wyniku błędów podczas replikacji
Część może powstawać w wyniku modyfikacji chemicznych

35
Jak może dojść do powstania substytucji
Formy tautomeryczne zasad azotowych adenina forma iminowa adeniny tymina forma enolowa tyminy

36
Formy tautomeryczne: iminowa i enolowa występują rzadko - 1: nukleotydów. Mogą (nie muszą) tworzyć pary niestandardowe:
 tworzyć pary niestandardowe:.")
37
enol - T – G enol - G – T imino - A – C imino - C – A
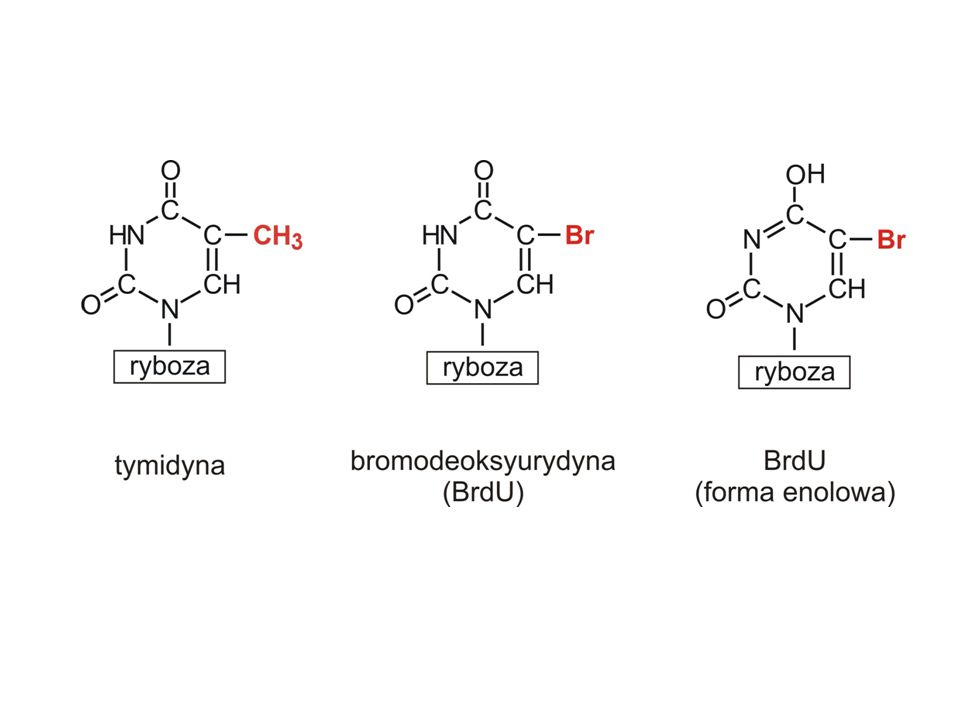
Niestandardowe pary zasad enol - T – G enol - G – T imino - A – C imino - C – A Karcynogen - 5 bromodeoksyurydyna (BrdU) jest analogiem tymidyny. Znacznie częściej niż tymidyna przyjmuje konformację enolową – częściej dochodzi do substytucji
 jest analogiem tymidyny. Znacznie częściej niż tymidyna przyjmuje konformację enolową – częściej dochodzi do substytucji.")
39
A – T A – BrdU enol-BrdU – G G – C podział 1 podział 2 podział 3
WNIOSEK: Tautomeryzacje mogą prowadzić do tranzycji

40
Przyjęcie konformacji „syn” przez purynę umożliwia zajście transwersji.
Może np. powstać para pomiędzy izomerami adenylanu: syn, amino - A : anty, imino - A

41
Polimeraza może popełniać błędy w wyniku „poślizgu na matrycy” (ang
Polimeraza może popełniać błędy w wyniku „poślizgu na matrycy” (ang. slippage errors). Może dodawać nukleotydy zwłaszcza w miejscach tandemowych powtórzeń W wyniku wypętlenia fragmentu DNA może dojść do pominięcia tego fragmentu przy replikacji (delecja). Szereg chorób o podłożu genetycznym jest związanych z wydłużaniem sekwencji repetytywnych (tandemowych). Klasyczny przykład: choroba Huntingtona (w białku huntingtinie trójka CAG kodująca glutaminę, powtórzeń – norma; powyżej 35 - choroba)
. Może dodawać nukleotydy zwłaszcza w miejscach tandemowych powtórzeń. W wyniku wypętlenia fragmentu DNA może dojść do pominięcia tego fragmentu przy replikacji (delecja). Szereg chorób o podłożu genetycznym jest związanych z wydłużaniem sekwencji repetytywnych (tandemowych). Klasyczny przykład: choroba Huntingtona (w białku huntingtinie trójka CAG kodująca glutaminę, powtórzeń – norma; powyżej 35 - choroba)")
42
Chemiczne modyfikacje zasad azotowych:
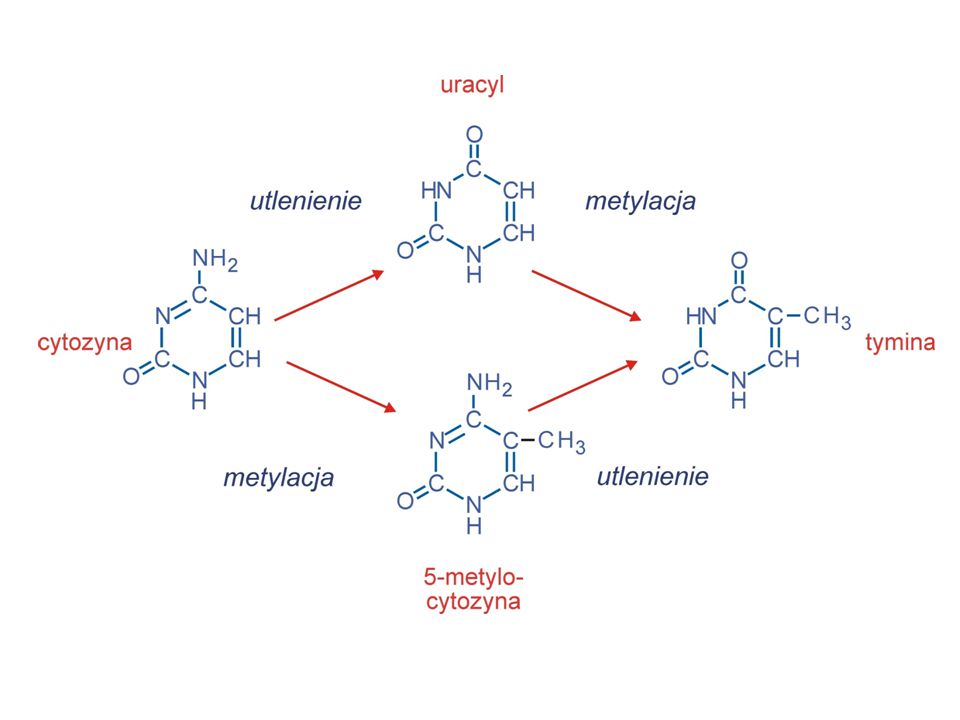
Utlenianie Alkilacja Utlenianie adeniny i cytozyny: NH =O cytozyna uracyl (może zachodzić w normalnych warunkach) adenina hipoksantyna (w DNA kompl. z C) Utlenienie grup aminowych adeniny i cytozyny może prowadzić do mutacji. guanina ksantyna (kompl. z C) Brak mutacji Typowym utleniaczem zasad azotowych jest HNO2.
 adenina hipoksantyna (w DNA kompl. z C) Utlenienie grup aminowych adeniny i cytozyny może prowadzić do mutacji. guanina ksantyna (kompl. z C) Brak mutacji. Typowym utleniaczem zasad azotowych jest HNO2.")
43
Utlenianie przez reaktywne formy tlenu (RFT) może prowadzić do utworzenia 8-oksyguanylanu.
8-oksyguanozyna Utlenienie guanylanu do 8-oksyguanylanu może prowadzić do transwersji: G T , gdyż 8-oxG może tworzyć parę z A.

44
Utlenianie C do U zachodzi dość często
Utlenianie C do U zachodzi dość często. W toku ewolucji wykształcił się specjalny mechanizm naprawy rozpoznający U w DNA. Może jednak dojść do metylacji U (do tyminy) i wówczas mutacja pozostaje niezauważona.
 i wówczas mutacja pozostaje niezauważona.")
45
U kręgowców i roślin główną metylotransferazą DNA jest enzym rozpoznający CpG i metylujący cytozynę.
Utlenianie 5-metylo-C do tyminy powoduje tranzycję C:G do T:A podczas kolejnych replikacji. Jest to najczęstsza mutacja u człowieka.

47
Podsumowanie: Utlenienie C do U - zauważone bo U jest nietypowe dla DNA – możliwość naprawy (chyba, że zanim U zostanie „odkryte” ulegnie metylacji) Utlenienie 5-metylo-C do T – niezauważone bo T jest typowe dla DNA – brak możliwości naprawy W wyniku tego, że DNA ulega metylacji przy C5 cytozyny i ze względu na to, że utlenienie 5-metylocytozyny pozostaje niezauważone, w toku ewolucji ilość par CpG w genomach kręgowców uległa znacznemu obniżeniu.

48
Częstość występowania sekwencji CpG:
Prokaryota: 1/16 nukleotydów (zgodnie z rachunkiem prawdopodobieństwa) Eukaryota: 1/50 nukleotydów Metylacja C w parach CpG: Prokaryota: 5% Eukaryota: % Ciekawostka: Dzięki temu - w toku ewolucji sekwencje oligonukleotydowe zawierające niemetylowane CpG stały się jednymi z sygnałów zagrożenia rozpoznawanymi przez układ odpornościowy (przez receptory TLR)
 Eukaryota: 1/50 nukleotydów. Metylacja C w parach CpG: Prokaryota: 5% Eukaryota: 70-90% Ciekawostka: Dzięki temu - w toku ewolucji sekwencje oligonukleotydowe zawierające niemetylowane CpG stały się jednymi z sygnałów zagrożenia rozpoznawanymi przez układ odpornościowy (przez receptory TLR)")
49
350 nukleotydów / 7 CpG = 50 350 nukleotydów / 29 CpG = 12 E. coli
CZŁOWIEK: GGCTTTTGCCTGGGGTTCTCAGGAGGGGAGAGTTGGGAGAGGCTTTGCTGCTGAGGAAATTTATTTGGTAGATTGAAGGTTTGAACGAGAGCTACAGAAACGAAAGAAAAAGTCTGTATAAGCCAATGGTGTTCGGGAAGAAAATAACCCCATTGCCTTGAGTTTGTAGGTGCCACTACTACTCTGGAAAAATGGCAGATGACGAAGACTATGAGGAGGTGGTGGAGTACTACACAGAAGAAGTGGTTTA CGAAGAGGTGCCGGGAGAGACAATAACAAAAATTTATGAGACTACGACAACAAGGACATCTGACTATGAGCAATCAGAAACTTCCAAACCAGCTCTGGCA 350 nukleotydów / 7 CpG = 50 E. coli TAGTTCTTAACGCCGATACCACATTCCATACCGTTACGGACTTCGTTAA CGTCATCTTTGAAGCGGAGCAGGGACTCCAGCTCGCCTTCGTAGATAACCACGTTGTCACGCAGAACGGGGATCGGGTTGTGACGTTTAACCACACCTTCGGTAACCATACAGCCTGCGATGGCACCAAATTTCGGCGATTTGAACACGTCACGAACTTCCGCCAGACCGATAATCTGCTGTTTCAGTTCCGGAGACAGCATACCGCTCATCGCTGCTTTCACTTCGTCAATCAGGTTATAGATGA CGGAGTAGTAACGCAGATCCAGGCTTTCCGCTTCAATCACTTTACGTGCA 350 nukleotydów / 29 CpG = 12

50
ZADANIE DOMOWE: Sprawdź czy podany przykład to reguła czy wyjątek Wybierz 3 dowolne (dość długie) fragmenty z genomu kręgowca – sprawdź ilość CpG Dla porównania – sprawdź częstość występowania CpG u bakterii

51
Czynniki alkilujące (dodające reszty metylowe lub etylowe)
N-metylo-nitrozoguanidyna N-metylo-N'-nitro-nitrosoguanidyna (MNNG) iperyt siarkowy iperyt azotowy metanosulfonian etylowy gaz bojowy od I wojny św. (Ypres 1917) Iperyt siarkowy Iperyt azotowy pochodne stosowane w onkologii – cyklofosfamid, ifosfamid
 iperyt siarkowy. iperyt azotowy. metanosulfonian etylowy. gaz bojowy. od I wojny św. (Ypres 1917) Iperyt siarkowy. Iperyt azotowy. pochodne stosowane w onkologii – cyklofosfamid, ifosfamid.")
52
Czynniki alkilujące metylują lub etylują zasady azotowe zaburzając strukturę helisy.
Przeniesienie grupy metylowej na O6 guaniny prowadzi do błędnego parowania zasad (O6-metyloguanina może ulegać parowaniu z tyminą). Przeniesienie grupy metylowej na N3 adeniny prowadzi do zatrzymania replikacji.
. Przeniesienie grupy metylowej na N3 adeniny prowadzi do zatrzymania replikacji.")
53
Pochodne iperytu azotowego alkilują dwa nukleotydy guanylowe przy N-7, wprowadzając wiązania krzyżowe do cząsteczki DNA (pomiędzy łańcuchami)
")
54
Związki o płaskiej strukturze zawierające kilka pierścieni mogą ulegać interkalacji pomiędzy zasady azotowe DNA i powodować insercje oraz delecje. Takie mutacje w obrębie sekwencji kodujących mogą zmieniać ramkę odczytu prowadząc do braku syntezy danego białka. benzopiren oranż akrydyny

55
Interkalacja bromku etydyny do DNA powoduje zmianę struktury DNA.
bromek etydyny Interkalacja bromku etydyny do DNA powoduje zmianę struktury DNA. EtBr służy do barwienia DNA w żelu. Można go obserwować w świetle UV.

56
Pod wpływem UV powstają dimery tyminy
Dimer tyminy blokuje replikację

57
Promieniowanie jonizujące
Może powodować drastyczne zmiany: Przerwanie wiązań fosfodiestrowych prowadzi do delecji Otwarcie pierścienia imidazolowego puryn usunięcie puryny przez glikozylazę powstanie miejsca apurynowego

58
NAPRAWA DNA Odwrócenie zmiany Wycięcie (NER, BER, MMR) Ad. 1
Fotoreaktywacja – w reakcji zależnej od światła (widzialnego) prowadzonej przez fotoliazę zostają zerwane wiązania w pierścieniu cyklobutanowym tworzącym dimer tyminy odtworzenie dwóch reszt tyminy. Fotoliaza (heterodimer kodowany przez geny phrA i phrB) występuje u bakterii i roślin, ale nie u ludzi. Usuwanie reszt metylowych przez enzym metylotransferazę O6‑metyloguaniny. Podczas reakcji ulega metylacji grupa –SH w centrum aktywnym enzymu – „mechanizm samobójczy”.
 prowadzonej przez fotoliazę zostają zerwane wiązania w pierścieniu cyklobutanowym tworzącym dimer tyminy odtworzenie dwóch reszt tyminy. Fotoliaza (heterodimer kodowany przez geny phrA i phrB) występuje u bakterii i roślin, ale nie u ludzi. Usuwanie reszt metylowych przez enzym metylotransferazę O6‑metyloguaniny. Podczas reakcji ulega metylacji grupa –SH w centrum aktywnym enzymu – „mechanizm samobójczy .")
59
Naprawa przez wycięcie (ang. excision repair)
Ad. 2 Naprawa przez wycięcie (ang. excision repair) NER – wycięcie nukleotydu (ang. nucleotide excision repair) BER – wycięcie zasady (ang. base excision repair) MMR – (ang. mismatch repair) Wszystkie polegają na trzyetapowym procesie: wycięcie uszkodzonego lub błędnego fragmentu, odtworzenie usuniętego fragmentu na matrycy DNA, ligacja
 NER – wycięcie nukleotydu (ang. nucleotide excision repair) BER – wycięcie zasady (ang. base excision repair) MMR – (ang. mismatch repair) Wszystkie polegają na trzyetapowym procesie: wycięcie uszkodzonego lub błędnego fragmentu, odtworzenie usuniętego fragmentu na matrycy DNA, ligacja.")
60
NER Mechanizm do usuwania dużych zmian powodujących wyraźne zaburzenie helisy – dimery tyminy, chemiczne addukty. Enzymy uczestniczące w tym procesie to ekscynukleazy (ang. excinucleases), ponieważ wycinają z nici DNA fragment zawierający błąd. U E. coli w proces NER zaangażowanych jest 5 białek: kompleks UvrABC rozpoznający uszkodzenie i usuwający fragment DNA, helikaza UvrD oraz polimeraza I. Białka Uvr odkryto na podstawie różnic między dzikim szczepem E. coli a mutantami wrażliwymi na promieniowanie UV (Uvr od ang. UV-resistance).
, ponieważ wycinają z nici DNA fragment zawierający błąd. U E. coli w proces NER zaangażowanych jest 5 białek: kompleks UvrABC rozpoznający uszkodzenie i usuwający fragment DNA, helikaza UvrD oraz polimeraza I. Białka Uvr odkryto na podstawie różnic między dzikim szczepem E. coli a mutantami wrażliwymi na promieniowanie UV (Uvr od ang. UV-resistance).")
61
NER wycięcie fragmentu przez UvrABC synteza DNA przez polimerazę DNA I
połączenie przez ligazę NER

62
Mechanizm naprawy zmian, które nie powodują lub powodują tylko niewielkie zaburzenie struktury DNA: np. usuwanie U lub utlenionych czy metylowanych zasad. BER Rozpoznawana jest nieprawidłowa struktura zasady. W pierwszym etapie hydrolizie ulega wiązanie glikozydowe pomiędzy nieprawidłową zasadą a deoksyrybozą. Proces prowadzą glikozylazy. Np. uracylo-N-glikozylaza rozpoznaje uracyl w DNA a glikozylaza metylopurynowa rozpoznaje metylowane G i A. Powstają miejsca apurynowe i apirymidynowe (miejsca AP) rozpoznawane przez endonukleazę AP, która przecina DNA po stronie 5’ miejsca AP. Fosfodiesteraza deoksyrybozowa wycina fosforan deoksyrybozy a polimeraza DNA I wprowadza brakujący nukleotyd.
 rozpoznawane przez endonukleazę AP, która przecina DNA po stronie 5’ miejsca AP. Fosfodiesteraza deoksyrybozowa wycina fosforan deoksyrybozy a polimeraza DNA I wprowadza brakujący nukleotyd.")
63
MMR Mechanizm naprawiający błędy powstałe podczas replikacji. Zwiększa dokładność replikacji, zmniejsza ilość błędów ok. 100 razy. Musi dojść do rozpoznania bardzo subtelnych różnic w strukturze DNA. Musi dojść do rozpoznania, który z dwóch nukleotydów w parze jest błędny!

64
MMR – c.d. Błędny jest ten występujący w nici świeżo spolimeryzowanej. Musi więc istnieć mechanizm rozróżniający nić matrycową od potomnej. Nić matrycowa jest rozpoznawana dzięki jej metylacji (np. u E. coli metylacja A w sekwencji GATC). Metylacja nowo utworzonej nici zachodzi w kilka minut po jej replikacji. Do czasu jej metylacji może zajść naprawa. U E. coli kompleks odpowiedzialny za MMR to MutHLS.
. Metylacja nowo utworzonej nici zachodzi w kilka minut po jej replikacji. Do czasu jej metylacji może zajść naprawa. U E. coli kompleks odpowiedzialny za MMR to MutHLS.")
65
Wiąże się do helisy; przyłącza się MutL (dimer).
Białko MutS rozpoznaje niekomplementarne pary nukleotydów wbudowane w DNA. Wiąże się do helisy; przyłącza się MutL (dimer).
.")
66
Białko MutH wiąże się do wszystkich hemimetylowanych sekwencji GATC.
Kompleks MutS-MutL przy udziale ATP przeciąga DNA aż do spotkania z najbliższymi MutH. Oddziaływanie MutH z MutL aktywuje endonukleazową aktywność MutH; MutH przecina niezmetylowaną nić DNA. Przy udziale UvrD (helikaza), egzonukleazy I i białek SSB dochodzi do usunięcia jednej i zabezpieczenia drugiej nici DNA. Powstała luka jest wypełniana przez kompleks polimerazy DNA III.
, egzonukleazy I i białek SSB dochodzi do usunięcia jednej i zabezpieczenia drugiej nici DNA. Powstała luka jest wypełniana przez kompleks polimerazy DNA III.")
67
Mutacje w genach kodujących białka odpowiedzialne za naprawę DNA prowadzą do groźnych chorób
Xeroderma pigmentosum (skóra pergaminowata, barwnikowa). Kilka postaci. Wszystkie związane z mutacjami w genach białek odpowiedzialnych za naprawę przez wycinanie. Najpowszechniej występująca – mutacje genów enzymów systemu NER.
. Kilka postaci. Wszystkie związane z mutacjami w genach białek odpowiedzialnych za naprawę przez wycinanie. Najpowszechniej występująca – mutacje genów enzymów systemu NER.")
68
skuteczna ochrona przed UV
Xeroderma pigmentosum skuteczna ochrona przed UV

69
Mutacje w obrębie genów kodujących białka uczestniczące w MMR towarzyszą rozwojowi nowotworu jelita grubego HNPCC HNPCC (ang. hereditary nonpolyposis colon cancer) U pacjentów stwierdzono mutacje w obrębie genu MLH1 (homolog MutL oraz genów MSH2 i MSH6 (homologi MutS).
 U pacjentów stwierdzono mutacje w obrębie genu MLH1 (homolog MutL oraz genów MSH2 i MSH6 (homologi MutS).")
Podobne prezentacje


















 organizmów eukariotycznych i większości wirusów posiada na swoim końcu 5nietypową strukturę zwaną>")
