Pobierz prezentację
Pobieranie prezentacji. Proszę czekać

1
TRANSLACJA BIOSYNTEZA BIAŁKA
przepisanie informacji zawartej w mRNA na cząsteczki efektorowe = powstanie łańcuchów polipeptydowych = utworzenie pierwszorzędowej struktury białek Biosynteza białka ogół procesów prowadzących do powstania NATYWNEJ struktury białka. obejmuje: translację – fałdowanie – potranslacyjne modyfikacje

2
Istotą procesu translacji jest tworzenie wiązań peptydowych
pomiędzy kolejnymi konkretnymi aminokwasami zgodnie z informacją zapisaną kodem genetycznym w genie kodującym dany łańcuch polipeptydowy

3
W procesie translacji uczestniczą CO? PO CO?
mRNA tRNA syntetazy aminoacylo-tRNA rybosomy aminokwasy czynniki białkowe matryca funkcja adaptorowa; konieczne, bo aminokwasy nie rozpoznają kodonów środowisko; enzym substraty regulatory

4
RYBOSOMY struktury o średnicy 20 nm, złożone zawsze z 2
podjednostek: dużej i małej obie podjednostki zbudowane są z rRNA i białek występują w każdej komórce (oprócz dojrzałych erytrocytów) są miejscem grupującym składniki uczestniczące w biosyntezie białka struktury stanowiące środowisko kontrolujące oddziaływania pomiędzy mRNA i aminoacylo-tRNA ogromne kompleksy enzymatyczne o złożonej strukturze prowadzące reakcję tworzenia wiązań peptydowych w oparciu o matrycę mRNA współdziałając z cytoplazmatycznymi, pomocniczymi czynnikami białkowymi umożliwiają zaistnienie całej gamy aktywności potrzebnych do wszystkich etapów translacji
 są miejscem grupującym składniki uczestniczące. w biosyntezie białka. struktury stanowiące środowisko kontrolujące. oddziaływania pomiędzy mRNA i aminoacylo-tRNA. ogromne kompleksy enzymatyczne o złożonej. strukturze prowadzące reakcję tworzenia wiązań. peptydowych w oparciu o matrycę mRNA. współdziałając z cytoplazmatycznymi, pomocniczymi czynnikami białkowymi umożliwiają. zaistnienie całej gamy aktywności potrzebnych do. wszystkich etapów translacji.")
5
Struktura małej podjednostki rybosomu Thermus thermophilus
Science 292 str , 2001 Crystal structure of the ribosome at 5.5 A resolution Yusupov MM, Yusupova GZ, Baucom A, Lieberman K, Earnest TN, Cate JH, Noller HF; University of California at Santa Cruz

6
Funkcjonowanie rybosomu
rRNA i białka rybosomów tworzą jedną interaktywną strukturę. rybosom posiada kilka miejsc aktywnych; każde z nich jest utworzone przez poszczególne grupy białek i odcinki rRNA. te grupy białek i odcinki rRNA zawsze funkcjonują w kontekście całego rybosomu.

7
tRNA są małe (73-93 nukleotydów; 25 kDa)
wszystkie tRNA mają podobną strukturę i szereg wspólnych cech stanowią 15% masy komórkowego RNA 10-25% nukleotydów zmodyfikowanych

8
KOD GENETYCZNY Phe Leu Ser Tyr Stop Cys Trp Pro His Gln Arg Ile Met
UUU UUC UUA UUG Phe Leu UCU UCC UCA UCG Ser UAU UAC UAA UAG Tyr Stop UGU UGC UGA UGG Cys Trp CUU CUC CUA CUG CCU CCC CCA CCG Pro CAU CAC CAA CAG His Gln CGU CGC CGA CGG Arg AUU AUC AUA AUG Ile Met ACU ACC ACA ACG Thr AAU AAC AAA AAG Asn Lys AGU AGC AGA AGG GUU GUC GUA GUG Val GCU GCC GCA GCG Ala GAU GAC GAA GAG Asp Glu GGU GGC GGA GGG Gly Nagroda Nobla z medycyny i fizjologii, 1968 Robert W. Holley, Har Gobind Khorana, Marshall W. Nirenberg „za rozszyfrowanie kodu genetycznego i wyjaśnienie jego funkcji w syntezie białka”

9
AMINOKWASY - KODONY – tRNA
Francis Crick – 1966 – hipoteza chwiejności zasad (wobble hypothesis) zasada 5’ antykodonu (tRNA) zasada 3’ kodonu (mRNA) A U A lub G C G U lub C I A lub U lub C
 zasada 5’ antykodonu (tRNA) zasada 3’ kodonu. (mRNA) A. U. A lub G. C. G. U lub C. I. A lub U lub C.")
10
Kodony dla seryny UCU, UCC, UCA, UCG AGU, AGC
Zgodnie z chwiejnością zasad: 5’ antykodonu (tRNA) 3’ kodonu (mRNA) A U A lub G C G U lub C I A lub U lub C Muszą istnieć trzy tRNA dla seryny. Potencjalne antykodony tRNA to (5’-3’): Albo: UGA, GGA i GCU Albo: IGA, CGA i GCU
 3’ kodonu (mRNA) A. U. A lub G. C. G. U lub C. I. A lub U lub C. Muszą istnieć trzy tRNA dla seryny. Potencjalne antykodony tRNA to (5’-3’): Albo: UGA, GGA i GCU. Albo: IGA, CGA i GCU.")
11
Działanie syntetaz aminoacylo-tRNA
Reakcja tworzenia aminoacylo-tRNA spełnia podwójną rolę: dopasowanie właściwego aminokwasu do właściwego tRNA zaktywowanie aminokwasu Reakcja jest dwustopniowa: I. PPi + H2O 2 Pi II. aminoacylo-AMP + tRNA aminoacylo-tRNA + AMP aminoacylo-AMP

12
Zapis sumaryczny aminokwas + ATP + tRNA + H2O
aminoacylo-tRNA + AMP + 2Pi

13
Syntetazy aminoacylo-tRNA ENZYMY O WYJĄTKOWO WYSOKIEJ SPECYFICZNOŚCI
Specyficznie rozpoznają dany aminokwas Specyficznie rozpoznają izoakceptorowe tRNA dla danego aminokwasu Syntetazy aminoacylo-tRNA posiadają „wewnętrzny mechanizm kontroli jakości” – centrum hydrolityczne, które hydrolizuje błędnie utworzone wiązania. SYNTETAZY STANOWIĄ JEDNEN Z ELEMENTÓW ODPOWIEDZIALNYCH ZA WIERNOŚĆ TRANSLACJI

14
Rybosom – tRNA – mRNA wzajemne wielkości mRNA 30 nukleotydów
10 kodonów mRNA E P A

15
MIEJSCA A, P, E w RYBOSOMIE
A - miejsce akceptorowe (aminokwasowe). W miejscu tym wyeksponowany jest kolejny, pusty jeszcze kodon odpowiadający kolejnemu aminokwasowi jaki ma się przyłączyć do łańcucha białka. Miejsce to wiąże wchodzący do rybosomu odpowiedni aa-tRNA. Jest tworzone przez małą i dużą podjednostkę rybosomu. P - miejsce peptydylowe Miejsce, w którym znajduje się peptydylo-tRNA. Jest tworzone przez małą i dużą podjednostkę rybosomu E – od ang. EXIT, wyjście Miejsce, przez które „pusty” tRNA opuszcza rybosom. Znajduje się w obrębie dużej podjednostki.
. W miejscu tym wyeksponowany jest kolejny, pusty jeszcze kodon odpowiadający kolejnemu aminokwasowi jaki ma się przyłączyć do łańcucha białka. Miejsce to wiąże wchodzący do rybosomu odpowiedni aa-tRNA. Jest tworzone przez małą i dużą podjednostkę rybosomu. P - miejsce peptydylowe. Miejsce, w którym znajduje się peptydylo-tRNA. Jest tworzone przez małą i dużą podjednostkę rybosomu. E – od ang. EXIT, wyjście. Miejsce, przez które „pusty tRNA opuszcza rybosom. Znajduje się w obrębie dużej podjednostki.")
16
Ogólny schemat przebiegu translacji

17
ETAPY TRANSLACJI INICJACJA obejmuje etapy przed utworzeniem pierwszego wiązania peptydowego. Obejmuje związanie mRNA do rybosomu oraz utworzenie kompleksu inicjującego. Jest to najwolniejszy etap translacji, decydujący o szybkości całej translacji i najsilniej poddany regulacji. ELONGACJA obejmuje reakcje od utworzenia pierwszego aż po ostatnie wiązanie peptydowe. TERMINACJA obejmuje uwolnienie polipeptydu z rybosomu oraz dysocjację rybosomów na podjednostki. Każdy z tych trzech etapów wymaga obecności specyficznych czynników cytoplazmatycznych. Energia na różnych etapach translacji jest dostarczana przez hydrolizę GTP.

18
Białka G pełnią funkcję
MOLEKULARNYCH PRZEŁĄCZNIKÓW (MOLECULAR SWITCH – ON / OFF) nie tylko w translacji GAP - GTPase Activating Protein (białko aktywujące GTPazę) GEF - Guanine nucleotide Exchange Factor (czynnik wymiany nukleotydów guaninowych) Alfred G. Gilman & Martin Rodbell, 1994 Nagroda Nobla z medycyny „for discovery of G-proteins and the role of these proteins in signal transduction in cells” (początki badań – 1980)
 nie tylko w translacji. GAP - GTPase Activating Protein. (białko aktywujące GTPazę) GEF - Guanine nucleotide Exchange Factor. (czynnik wymiany nukleotydów guaninowych) Alfred G. Gilman & Martin Rodbell, 1994 Nagroda Nobla z medycyny. „for discovery of G-proteins and the role of these proteins in signal transduction in cells (początki badań – 1980)")
19
WSPÓLNE DLA PROKARYOTA I EUKARYOTA ETAPY INICJACJI TRANSLACJI
mała podjednostka rybosomu wiąże inicjatorowy tRNA - ten, który niesie pierwszy aminokwas łańcucha polipeptydowego mała podjednostka rybosomu jest rekrutowana do mRNA i umieszczana w miejscu kodonu inicjacji (initiation codon) przyłącza się duża podjednostka – tworzy się cały RYBOSOM
 przyłącza się duża podjednostka – tworzy się cały RYBOSOM.")
20
(INITIATION FACTORS, IF)
CZYNNIKI INICJUJĄCE (INITIATION FACTORS, IF) Są to białkowe czynniki uczestniczące tylko w inicjacji. Wiążą się tylko z małą podjednostką rybosomu i tylko do czasu jej asocjacji z dużą podjednostką. Wtedy oddysocjowują. NIE MA ICH W RYBOSOMIE 70S (80S) Ta aktywność odróżnia IF od strukturalnych (integralnych) białek rybosomów. Prokariotyczne czynniki inicjujące – skrót IF Eukariotyczne czynniki inicjujące oznaczamy eIF (eukaryotic initiation factors)
 Są to białkowe czynniki uczestniczące tylko w. inicjacji. Wiążą się tylko z małą podjednostką rybosomu. i tylko do czasu jej asocjacji z dużą. podjednostką. Wtedy oddysocjowują. NIE MA ICH W RYBOSOMIE 70S (80S) Ta aktywność odróżnia IF od strukturalnych. (integralnych) białek rybosomów. Prokariotyczne czynniki inicjujące – skrót IF. Eukariotyczne czynniki inicjujące oznaczamy. eIF (eukaryotic initiation factors)")
21
PROKARIOTYCZNE CZYNNIKI INICJUJĄCE
IF IF IF-3 IF-1 - jego rola jest najsłabiej poznana. Wiąże się do podjednostki 30S tylko jako część kompleksu inicjacyjnego. Zasłania miejsce A rybosomu. IF-2 - wiąże specyficznie inicjatorowy tRNA i kontroluje jego wejście do rybosomu. Do swej aktywności IF-2 musi być związany z GTP. Jest białkiem G. IF-3 - odgrywa podwójną rolę: 1. Utrzymuje pewną pulę podjednostek 30S w stanie wolnym. Uniemożliwia ich reasocjację z 50S. 2. Kontroluje zdolność 30S do związania mRNA. 30S musi związać IF-3, aby utworzyć kompleks inicjacyjny z mRNA.

22
INICJACJA

23
Kolejność zdarzeń (hipoteza)
Mała podjednostka wiąże IF-3 Rekrutacja mRNA Wiązanie IF2-fMet-tRNA w miejscu P rybosomu Związanie IF2-fMet-tRNA do miejsca P rybosomu zmienia konformację w IF2 i stymuluje wymianę GDP GTP Obecność GTP w IF2 sprzyja tworzeniu kompletnego rybosomu (70S). Następuje przyłączenie dużej podjednostki rybosomu Hydroliza GTP przy udziale białek L7/L12 dużej podjednostki rybosomu i uwolnienie czynników inicjujących z rybosomu.
. Następuje przyłączenie dużej podjednostki rybosomu. Hydroliza GTP przy udziale białek L7/L12 dużej podjednostki rybosomu i uwolnienie czynników inicjujących z rybosomu.")
24
Na czym polega aktywująca rola GTP w przypadku IF2?
Obecność GTP determinuje taką konformację IF2, która ułatwia przyłączenie dużej podjednostki rybosomu. Hydroliza GTP do GDP powoduje zmianę konformacji i zmniejszenie powinowactwa IF2 do rybosomu Funkcję czynnika GAP dla IF2-GTP pełni białko L7/L12.

25
INICJATOROWY tRNA Synteza białka zaczyna się od metioniny - metionina jest pierwszym aminokwasem tworzącego się łańcucha polipeptydowego Sygnał stanowi inicjatorowy kodon AUG Istnieją dwa typy tRNA mogących wiązać ten aminokwas i ten kodon (AUG): Uczestniczący w elongacji: Uczestniczący w inicjacji: Ta sama syntetaza aminoacylo-tRNA prowadzi obie reakcje (przyłącza resztę metioniny do obu rodzajów tRNA U bakterii oraz w organellach (chloroplastach, mitochondriach) inicjatorowy tRNA wiąże metioninę, której grupa aminowa zostaje później FORMYLOWANA N-formylo-metionylo-tRNA (fMet-tRNAf).
: Uczestniczący w elongacji: Uczestniczący w inicjacji: Ta sama syntetaza aminoacylo-tRNA prowadzi obie reakcje (przyłącza resztę metioniny do obu rodzajów tRNA. U bakterii oraz w organellach (chloroplastach, mitochondriach) inicjatorowy tRNA wiąże metioninę, której grupa aminowa zostaje później FORMYLOWANA N-formylo-metionylo-tRNA (fMet-tRNAf).")
26
fmet-tRNAf nie może przyłączyć się do miejsca A rybosomu.
Dlaczego istnieją różne tRNA dla pierwszej metioniny i pozostałych reszt metioniny? fmet-tRNAf nie może przyłączyć się do miejsca A rybosomu. met-tRNAm nie może przyłączyć się do miejsca P rybosomu

27
Inicjacja u bakterii wymaga oddziaływania pomiędzy mRNA i rRNA
Skąd rybosom „wie”, który kodon AUG jest startowy? E. coli ’ CCUCCUUA 3’ Salmonella typhimurium ’ CCUCCUUA 3’ Bordetella holmesii ’ CCUCCU 3’

28
5'-AGGAGG-3' sekwencja Shine-Dalgarno
W obrębie ok.10 nukleotydów powyżej miejsca startu znajduje się heksamer lub przynajmniej jego część: 5'-AGGAGG-3' fragment polipurynowy określany jako sekwencja Shine-Dalgarno

29
U Eukaryota nie ma takiego mechanizmu
U Eukaryota rybosom za pośrednictwem białek wiąże się z czapeczką, skanuje 5’UTR i rozpoczyna translację od pierwszego AUG (w >90% przypadków). Rozpoznawanie właściwego kodonu AUG jest wspomagane przez sekwencję sąsiadujących z nim nukleotydów. Optymalna sekwencja to: G C C A(G) C C AUG G Najważniejsze są: puryna o 3 nukleotydy przed AUG i nukleotyd guanylowy tuż za trójką AUG. Wtedy inicjacja translacji jest najwydajniejsza. Ta sekwencja to tzw. sekwencja Kozak (Kozak sequence; Kozak consensus sequence)
. Rozpoznawanie właściwego kodonu AUG jest wspomagane przez sekwencję sąsiadujących z nim nukleotydów. Optymalna sekwencja to: G C C A(G) C C AUG G. Najważniejsze są: puryna o 3 nukleotydy przed AUG i nukleotyd guanylowy tuż za trójką AUG. Wtedy inicjacja translacji jest najwydajniejsza. Ta sekwencja to tzw. sekwencja Kozak. (Kozak sequence; Kozak consensus sequence)")
30
Etapy inicjacji translacji u Eukaryota
Utworzenie kompleksu preinicjacyjnego 43S (mała podjednostka rybosomu + niektóre czynniki inicjacyjne + Met-tRNAi) Rekrutacja kompleksu 43S do czapeczki mRNA Skanowanie 5’UTR i rozpoznanie kodonu startowego Przyłączenie dużej podjednostki rybosomu
 Rekrutacja kompleksu 43S do czapeczki. mRNA. Skanowanie 5’UTR i rozpoznanie kodonu. startowego. Przyłączenie dużej podjednostki rybosomu.")
31
Aktywacja czynnika eIF2 przez wymianę GDP na GTP:
Związanie GTP przez eIF2 zwiększa jego powinowactwo do Met-tRNAi, eIF2 wiąże Met-tRNAi. Powstaje kompleks: eIF2GTP Met-tRNAi Ten kompleks wiąże się do 40S w miejscu P. Powstanie kompleksu eIF2-GTP-Met-tRNAi jest konieczne do tego, aby mała podjednostka związała się z mRNA.

32
Mała podjednostka rybosomu oddziałuje z mRNA za pośrednictwem białek:
cap-mRNA Białko Białko 40S eIF4E (25 kDa) wiąże się z czapeczką. eIF4E = białko wiążące czapeczkę (mRNA-cap binding protein) eIF4G (154 kDa) - rusztowanie eIF4A (46 kDa) - helikaza
 wiąże się z czapeczką. eIF4E = białko wiążące czapeczkę. (mRNA-cap binding protein) eIF4G (154 kDa) - rusztowanie. eIF4A (46 kDa) - helikaza")
33
N-końcowa domena czynnika eIF4G oddziałuje z białkiem wiążącym czapeczkę.
C-końcowa domena czynnika eIF4G oddziałuje z czynnikiem eIF3.

34
Skanowanie 5’UTR i rozpoznanie kodonu startowego

35
Te zmiany konformacyjne są związane z funkcjonowaniem białek G.
Po rozpoznaniu kodonu AUG przez antykodon inicjatorowego tRNA następują zmiany konformacyjne w obrębie kilku czynników inicjatorowych prowadzące do przyłączenia dużej podjednostki rybosomu. Te zmiany konformacyjne są związane z funkcjonowaniem białek G.

36
Regulacja szybkości przebiegu translacji
Zachodzi głównie na etapie INICJACJI (jest to korzystne energetycznie). Regulacja może dotyczyć: całego procesu inicjacji (wszystkich białek) pojedynczych, poszczególnych białek W regulacji uczestniczą: elementy cis-regulatorowe (znajdujące się w obrębie cząsteczki poddanej regulacji) elementy trans-regulatorowe (pochodzące spoza cząsteczki podlegającej regulacji) ELEMENTY CIS (tu wymienione elementy „samodzielne”) sekwencje i struktury otaczające kodon inicjatorowy struktura 5'UTR (skanowanie, współzawodnictwo między różnymi mRNA o czynniki inicjatorowe)
. Regulacja może dotyczyć: całego procesu inicjacji (wszystkich białek) pojedynczych, poszczególnych białek. W regulacji uczestniczą: elementy cis-regulatorowe (znajdujące się w obrębie cząsteczki poddanej regulacji) elementy trans-regulatorowe (pochodzące spoza cząsteczki podlegającej regulacji) ELEMENTY CIS (tu wymienione elementy „samodzielne ) sekwencje i struktury otaczające kodon. inicjatorowy. struktura 5 UTR (skanowanie, współzawodnictwo między różnymi mRNA o. czynniki inicjatorowe)")
37
Szybkość translacji niektórych białek zależy od poziomu żelaza w komórce
Ferrytyna – białko komórkowe magazynujące żelazo (4500 Fe / 1 cząsteczkę białka). Gdy w komórce brakuje żelaza, poziom ferrytyny może ulec obniżeniu. Fe - translacja mRNA ferrytyny W 5’ UTR mRNA ferrytyny znaleziono charakterystyczną strukturę/sekwencję nazwaną IRE (Iron Responsive Element)
. Gdy w komórce brakuje żelaza, poziom ferrytyny może ulec obniżeniu. Fe - translacja mRNA ferrytyny W 5’ UTR mRNA ferrytyny znaleziono charakterystyczną strukturę/sekwencję nazwaną IRE (Iron Responsive Element)")
38
IRE może przyłączać z wysokim powinowactwem IRP - Iron Regulatory Proteins (np. IRP1)
IRE – elementy cis-regulatorowe IRP – elementy trans-regulatorowe Kompleksy IRE/IRP stanowią przeszkodę przy skanowaniu 5’UTR przez małą podjednostkę rybosomu. IRP1 to białko żelazo-siarkowe.

39
Mechanizm działania IRP1
Fe IRP-1 pozostaje związane z żelazem (z centrami żelazo-siarkowymi) i nie oddziałuje z IRE. Zachodzi bez przeszkód translacja mRNA ferrytyny. ferrytyna
 i nie oddziałuje z IRE. Zachodzi bez przeszkód translacja mRNA ferrytyny. ferrytyna ")
40
Prokaryota EF-Tu EF-Ts EF- G Eukaryota eEF-1A eEF-1B eEF-2
ELONGACJA Rozpoczyna się od wprowadzenia do rybosomu drugiego aminoacylo-tRNA Kończy się z chwilą osiągnięcia przez rybosom kodonu „STOP”. Istota elongacji polega na tworzeniu wiązań peptydowych i translokacji rybosomu względem mRNA. Przebiega podobnie u Prokaryota i Eukaryota. Uczestniczą w niej cytoplazmatyczne czynniki elongacyjne: Prokaryota EF-Tu EF-Ts EF- G Eukaryota eEF-1A eEF-1B eEF-2

41
Porównanie IF2 i EF-Tu IF-2 EF-Tu
wprowadza inicjatorowy aminoacylo-tRNA do miejsca P rybosomu wprowadza aminoacylo-tRNA do miejsca A rybosomu wiąże GTP lub GDP IF2GTP – aktywny EF-TuGTP – aktywny tworzy kompleks IF2GTPfMet-tRNAf tworzy kompleks EF-TuGTPaa-tRNA

42
EF-Tu jest białkiem G EF-Tu•GTP - aktywny –
L7/L12 EF-Tu•GTP - aktywny – ma wysokie powinowactwo do aa-tRNA oraz do miejsca A rybosomu EF-Tu •GDP - nieaktywny – ma niskie powinowactwo do aa-tRNA oraz do miejsca A rybosomu

43
FUNKCJONOWANIE EF-Tu i EF-Ts
EF-Tu tworzy kompleks EF-TuGTPaa-tRNA i wprowadza aa-tRNA do miejsca A rybosomu. Ten kompleks może oddziaływać tylko z miejscem A takiego rybosomu, którego miejsce P jest zajęte. Oddziaływanie kodon-antykodon powoduje zmianę konformacyjną w obrębie EF-Tu umożliwiającą hydrolizę GTP GDP. Kompleks EF-TuGDP ma niskie powinowactwo do aa-tRNA i oddysocjowuje. Opuszcza rybosom. EF-Ts pełni funkcję GEF dla EF-Tu. EF-Ts wypiera GDP z cząsteczki EF-Tu. Tworzy się kompleks EF-TuEF-Ts. EF-Ts jest w tym kompleksie zastępowany przez GTP odtworzenie aktywnego EF-TuGTP. EF-TuGTP natychmiast wiąże się z kolejnym aa-tRNA i wprowadza go do miejsca A rybosomu.

44
DOŚWIADCZENIA: Stosując różne inhibitory stwierdzono, że: GTP związany z EF-Tu musi ulec hydrolizie do GDP, aby EF-Tu mógł odłączyć się od rybosomu Aby powstało wiązanie peptydowe EF-Tu musi opuścić rybosom

45
KONTROLA JAKOŚCI To, że obecność EF-Tu hamuje tworzenie wiązania peptydowego jest NIEZWYKLE WAŻNE stanowi mechanizm decydujący o bezbłędności translacji. Konformacja miejsca A pasuje do każdego EF-TuGTPaa-tRNA do miejsca A wejść może każdy aa-tRNA. Pozostać powinien tylko taki, którego antykodon pasuje do kodonu. Jeśli wejdzie nieprawidłowy aa-tRNA to brak lub bardzo słabe oddziaływanie kodon/antykodon powoduje, że nieprawidłowy aa-tRNA oddysocjowuje z miejsca A. Musi to nastąpić zanim zostanie utworzone wiązanie peptydowe. Podczas translacji nie zachodzi korekta (proof-reading) charakterystyczna dla procesu replikacji wbudowywanie aminokwasu musi być od razu prawidłowe. Fakt, że obecność EF-Tu uniemożliwia utworzenie wiązania peptydowego daje CZAS na to, aby nieprawidłowy aa-tRNA oddysocjował z miejsca A rybosomu. Błędnie wprowadzony aminokwas: 1/
 charakterystyczna dla procesu replikacji wbudowywanie aminokwasu musi być od razu prawidłowe. Fakt, że obecność EF-Tu uniemożliwia utworzenie wiązania peptydowego daje CZAS na to, aby nieprawidłowy aa-tRNA oddysocjował z miejsca A rybosomu. Błędnie wprowadzony aminokwas: 1/")
46
UTWORZENIE WIĄZANIA PEPTYDOWEGO
Aktywność odpowiedzialna za tę reakcję została nazwana: aktywnością peptydylotransferazy. Aktywność peptydylotransferazy znajduje się w dużej podjednostce rybosomu, w miejscu gdzie znajdują się blisko siebie końce aa-tRNA i peptydylo-tRNA. Zarówno białka tego obszaru rybosomu, jak i rRNA są konieczne dla tej aktywności. rRNA tworzy centrum katalityczne peptydylotransferazy. Utworzenie wiązania peptydowego nie wymaga na tym etapie dostarczenia energii. Energia wykorzystywana do powstania wiązania peptydowego została już wcześniej zmagazynowana w wiązaniu estrowym między resztą karboksylową aminokwasu a resztą hydroksylową końcowego nukleotydu adeninowego tRNA.

47
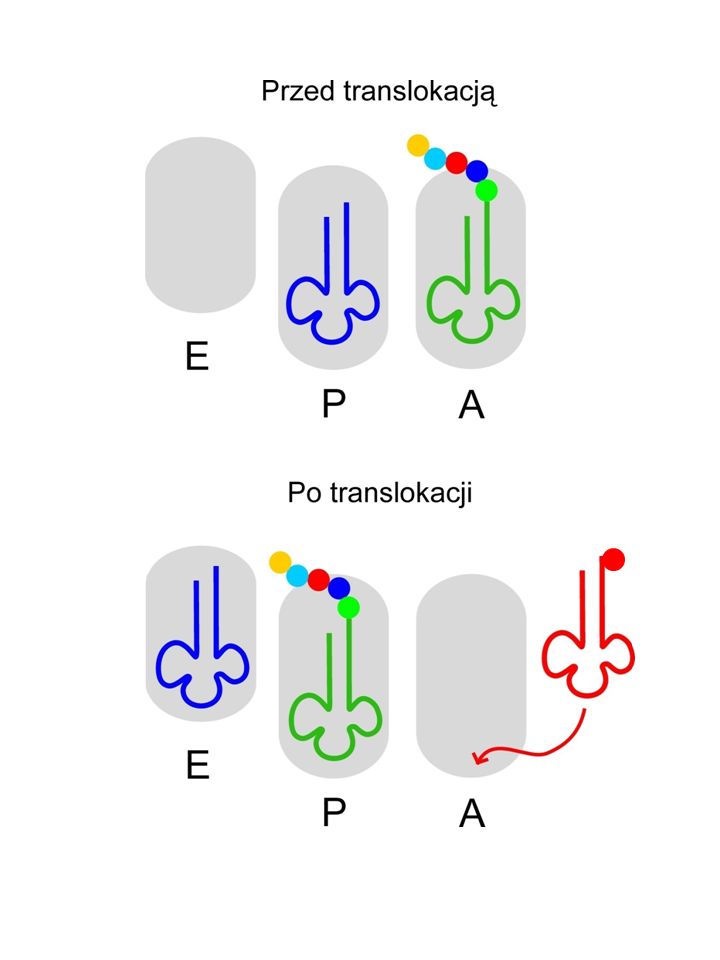
TRANSLOKACJA Następuje po utworzeniu wiązania peptydowego
W wyniku translokacji deacylowany tRNA przenosi się do miejsca E. Peptydylo-tRNA przenosi się z miejsca A do miejsca P. Miejsce A zostaje wolne dla następnego aa-tRNA.

49
Jeden z modeli translokacji
Tu uczestniczy czynnik EF-G Według tej koncepcji tworzący się peptyd w zasadzie nie opuszcza miejsca P w dużej podjednostce.

50
Rybosomy nie mogą wiązać równocześnie
EF-Tu i EF-G, a więc elongacja następuje zgodnie z cyklicznymi zmianami obecności tych czynników w rybosomie.

51
TERMINACJA Terminacja translacji następuje wtedy, gdy
w miejscu A rybosomu znajdzie się jeden z kodonów terminacyjnych (kodonów STOP): UAG (amber) UAA (ochre) UGA (opal)
: UAG (amber) UAA (ochre) UGA (opal)")
52
RF-1 - rozpoznaje UAG i UAA RF-2 - rozpoznaje UGA i UAA
CZYNNIKI UWALNIAJĄCE (RELEASE FACTORS) Nie istnieją (fizjologicznie) tRNA, które miałyby antykodony komplementarne do kodonów STOP jeśli w miejscu A rybosomu znajdzie się kodon STOP – żaden tRNA nie oddziałuje z miejscem A. Zakończenie translacji wymaga 2 klas białkowych czynników uwalniających: Czynniki uwalniające zależne od kodonu niezależne od kodonu Prokaryota RF-1, RF-2 RF-3 Eukaryota eRF-1 eRF-3 RF-1 - rozpoznaje UAG i UAA RF-2 - rozpoznaje UGA i UAA
 Nie istnieją (fizjologicznie) tRNA, które miałyby antykodony komplementarne do kodonów STOP jeśli w miejscu A rybosomu znajdzie się kodon STOP – żaden tRNA nie oddziałuje z miejscem A. Zakończenie translacji wymaga 2 klas białkowych czynników uwalniających: Czynniki uwalniające. zależne. od kodonu. niezależne od kodonu. Prokaryota. RF-1, RF-2. RF-3. Eukaryota. eRF-1. eRF-3. RF-1 - rozpoznaje UAG i UAA. RF-2 - rozpoznaje UGA i UAA.")
53
Należy zwrócić uwagę na mimikrę pomiędzy kwasem nukleinowym i białkiem

55
Terminacja – etapy Hydroliza wiązania estrowego między tRNA a łańcuchem polipeptydowym (uwolnienie białka) Oddysocjowanie czynników terminacji (funkcjonowanie białka G (RF3)) Oddysocjowanie deacylowanych tRNA Rozłączenie podjednostek rybosomu
) Oddysocjowanie deacylowanych tRNA. Rozłączenie podjednostek rybosomu.")
Podobne prezentacje









, jest jednym z 20 aminokwasów.>")










